 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 5, August 2005, pp. 495-500 Identification of Anopheles (Nyssorhynchus) albitarsis complex species (Diptera: Culicidae) using rDNA internal transcribed spacer 2-based polymerase chain reaction primes Cong Li, Richard C Wilkerson+ Walter Reed Army Institute of Research, Dept. of Entomology, 503 Robert Grant Ave., Silver Spring, MD 20910-7500, US Financial support:
NIH AI RO154139 to Jan Conn This research was performed under a Memorandum of Understanding between the Walter Reed Army Institute of Research and the Smithsonian Institution, with institutional support provided by both organizations. The material to be published reflects the views of the authors and should not be construed to represent those of the Department of the Army or the Department of Defense. Received 24 January
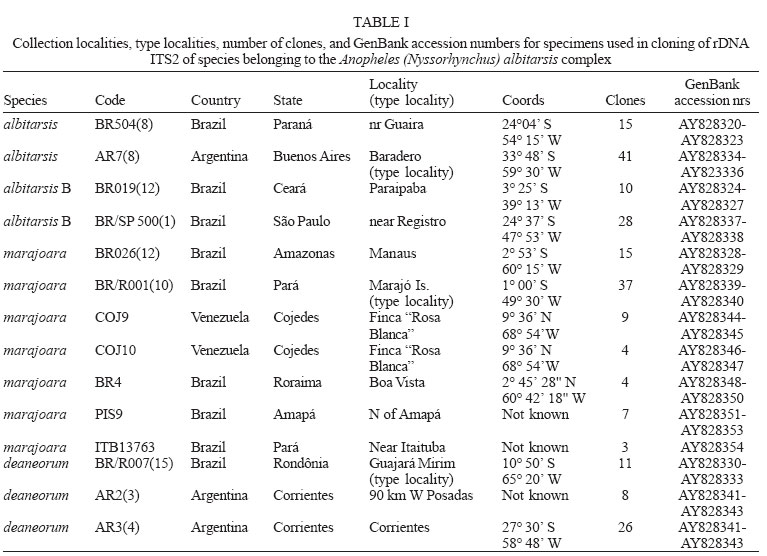
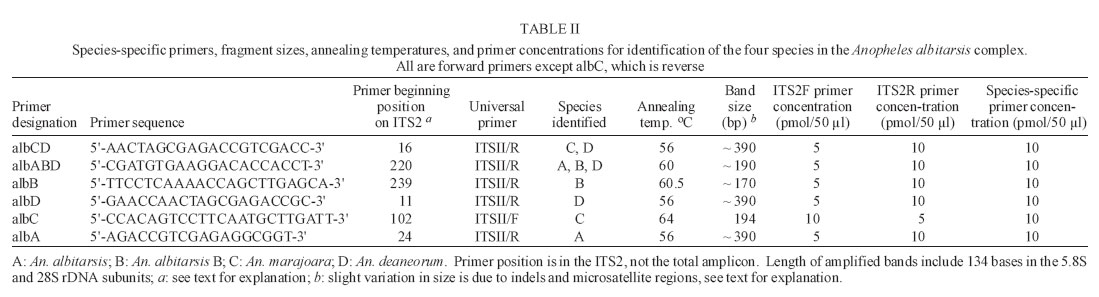
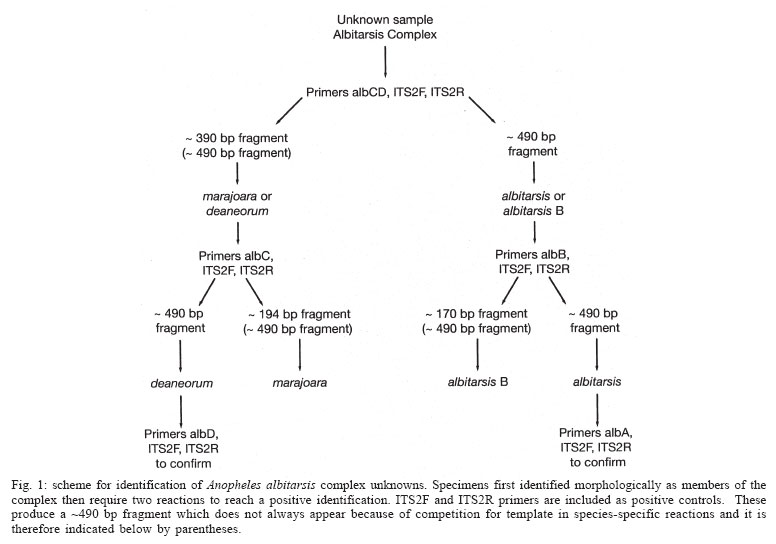
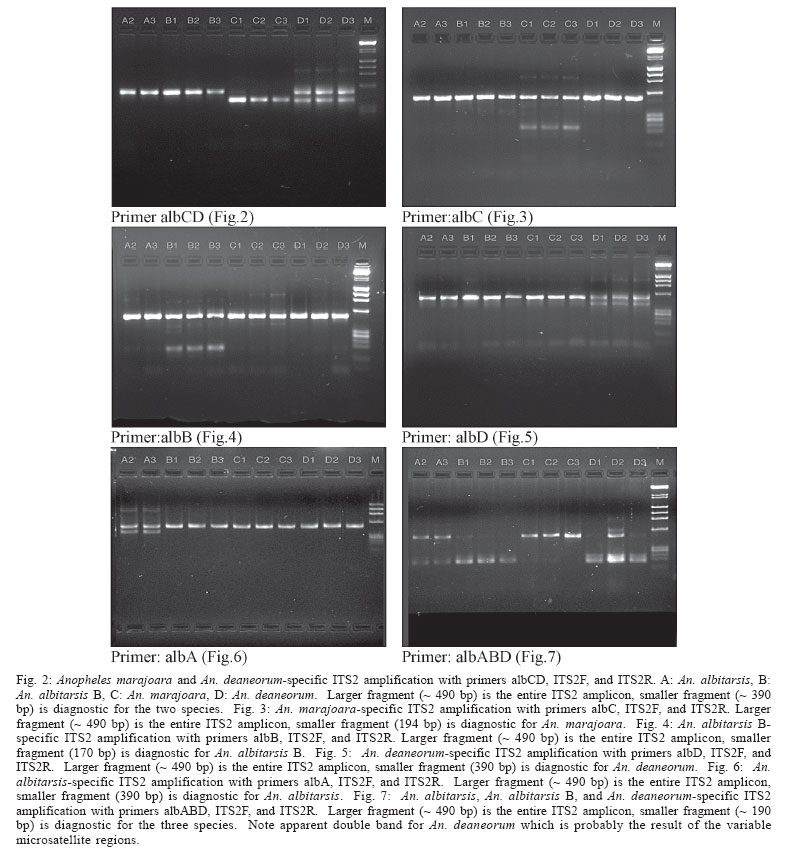
2005 Code Number: oc05121 Anopheles (Nyssorhynchus) marajoara is a proven primary vector of malaria parasites in Northeast Brazil, and An. deaneorum is a suspected vector in Western Brazil. Both are members of the morphologically similar Albitarsis Complex, which also includes An. albitarsis and an undescribed species, An. albitarsis "B". These four species were recognized and can be identified using random amplified polymorphic DNA (RAPD) markers, but various other methodologies also point to multiple species under the name An. albitarsis. We describe here a technique for identification of these species employing polymerase chain reaction (PCR) primers based on ribosomal DNA internal transcribed spacer 2 (rDNA ITS2) sequence. Since this method is based on known sequence it is simpler than the sometimes problematical RAPD-PCR. Primers were tested on samples previously identified using RAPD markers with complete correlation. Key words: Anopheles albitarsis - species complex - polymerase chain reaction identification - malaria - internal transcribed spacer 2 The Neotropical Anopheles (Nyssorhynchus) al-bitarsis complex currently consists of four morphologically similar species, at least one of which, An. marajoara Galvão & Damasceno, is an important malaria vector in the state of Amapá, Brazil (Conn et al. 2002). Prior to Linthicum's (1988) revision of the Argyritarsis Section, which includes the Albitarsis Complex, the only species of the four currently recognized species considered valid was An. albitarsis Lynch-Arribalzaga. Linthicum (1988) resurrected a second species, An. marajoara, from synonymy and reviewed the taxonomic history of An. albitarsis and related species. He also synonymized An. domesticus Galvão & Damesceno with An. marajoara and declared An. allopha Peryassu to be a nomen dubium. Subsequently, Rosa-Freitas (1989) described a third species, An. deaneorum, also a potential malaria vector in the state of Rondônia in Western Brazil (Klein et al. 1991a,b), and Wilkerson et al. (1995a,b), using random amplified polymorphic DNA-polymerase chain reaction (RAPD-PCR), discovered another as yet undescribed species, An. albitarsis species "B". Many suspected An. albitarsis s.l. to be a species complex because of its wide distribution and great morphological and behavioral variability. Various methods were used to make sense of the An. albitarsis puzzle, including chromosomes, allozymes, mitochondrial DNA restriction fragment length polymorphisms (mtDNA RFLPs), RAPD-PCR, morphology, and behavior (Kreutzer et al. 1976, Rosa-Freitas & Deane 1989, Rosa-Freitas et al. 1990, Narang et al. 1993, Wilkerson et al. 1995a,b). Wilkerson et al. (1995a,b), using RAPD-PCR, demonstrated the existence of the four currently recognized species (but see Lehr et al. 2005) and found 2-4 diagnostic RAPD primers for each that serve to identify individuals from throughout their ranges. RAPD-PCR has been the only method to discriminate among the four with certainty since only An. deaneorum can be reliably separated morphologically, and then only in the larval stage. The species are distributed in Central and South America as follow: An. albitarsis (Southern Brazil, Northern Argentina, Paraguay), An. deaneorum (Northern Argentina to Western Brazil), An. marajoara (Brazil, Venezuela, Colombia, Southern Central America), and An. albitarsis B (South, Central, and Eastern Brazil). While reproducible in any given lab that has standardized the technique, RAPD-PCR can be unreliable as an identification method when differing extraction protocols, reagents, and PCR machines are used. In addition, the above method (Wilkerson et al. 1995a,b) can require two or more reactions to make an identification. Because of the medical importance of the complex we sought a means, other than RAPD-PCR, to carry out identifications in support of epidemiological, ecological, and population genetics studies. With the above objective in mind, we investigated the rDNA ITS2 for species-specific sequence variation. Ribosomal DNA has been utilized for systematics and phylogenetics studies in a wide variety of organisms (e.g. Hillis & Dixon 1991, Miller et al. 1997 for Diptera). The functional regions that produce the ribosomes are highly conserved, but the transcribed (ITS1 and ITS2) and non-transcribed spacer (intergenic spacer, IGS) regions have high interspecific and low intraspecific variability, making them useful for explaining relationships of recently diverged species, and also useful as a basis for PCR identification of morphologically similar species. ITS2 sequences usually allow unambiguous species identifications in closely related Anopheles species (reviewed by Collins & Paskewitz 1996). Other examples since the above review include: An. (Ano.) quadrimaculatus complex (Cornel et al. 1996); An. (Cellia) maculatus complex (Torres et al. 2000); An. (Cel.) dirus complex (Walton et al. 1999); An. (Cel.) fluviatilis complex (Manonmani et al. 2001); An. (Ano.) bancroftii group (Beebe et al. 2001); An. (Cel.) punctulatus group (Beebe et al. 1999); An. (Cel.) funestus and related species (Hackett et al. 2000); and, An. (Ano.) crucians complex (Wilkerson et al. 2004). Rarely, the ITS2 sequences among cryptic species are too close to allow PCR-based identification. The only published example is An. (Ano.) inundatus Reinert (as C2) and An. (Ano.) diluvialis Reinert (as C1) of the An. quadrimaculatus complex (Cornel et al. 1996). We describe here development of rDNA ITS2 primers based on cloned sequence (Li & Wilkerson, unpublished data). This technique makes possible reliable identifications for those studying ecology, behavior and malaria transmission in relation to these species. MATERIALS AND METHODS Source of specimens - Specimens used for testing of PCR primers are the same as those reported in Wilkerson et al. (1995a,b). These represent samples of the four species from widely separated localities as described in the introduction and include the type localities for the three named species. Additional specimens from subsequent studies were also tested from Venezuela (Rubio-Palis et al. 2004), Trinidad, Boa Vista (state of Roraima, Brazil) and localities in the state of Amapá, Brazil. Localities and coordinates can be found in Table I and in the above references. Cloning - Locality and other information for specimens used to obtain clones, as well as Genbank accession numbers, are given in Table I (unpublished data, Li and Wilkerson). These clones are the basis for the species-specific primers described here. Detailed findings will be published elsewhere. Species identification - Peyton et al. (1992) provide a definition of An. subgenus Nyssorhynchus. Adult female Nyssorhynchus mosquitoes were first placed in the Albitarsis Complex by the following combination of characters (Linthicum 1988): hindtarsomere 5 without a basal dark band, vein 1A mostly pale-scaled (Argyritarsis Section), sternum I with a distinct row of white scales on each side and, posterolateral scale tufts beginning on segments III or IV. The presumed sister species to the Albitarsis Complex, An. braziliensis (Chagas), has the rows of white scales on sternum I but the posterolateral tufts begin on segment II. Additional An. braziliensis differences can also be found in the adult female, male genitalia, and pupal and larval stages (Linthicum 1988). Species determinations were then made using RAPD-PCR as described in Wilkerson et al. (1995a,b) and corroborated with the primers described here. DNA isolation - DNA was isolated from individual adult mosquitoes by phenol-chloroform extraction as described in Wilkerson et al. (1993). PCR amplification - Conserved primers flanking the ITS2 region correspond to conserved sequences in the 5.8S and 28S ribosomal subunits of An. quadrimaculatus complex species (Cornel et al. 1996): ITS2F 5'-TGTGAACTGCAGGACACATGAA-3' (forward) and ITS2R 5'-ATGCTTAAATTTAGGGGGTAGTC-3'(reverse). These, in conjunction with single species-specific primers (Table II), designed from cloned sequence (Table I), were used to identify the species. Fragment sizes listed in Table II include the ITS2 (Cornel et al. 1996) and 134 base pairs from the flanking 5.8S and 28S rDNA subunits. Primer beginning position numbers shown in Table II refer to ITS2 sequence (without flanking sequence) as reported by Li & Wilkerson in Genbank (Table I). Reactions were carried out in a total volume of 50 µl using the PCR buffer (GeneAmp 10´ Buffer II) supplied with AmpliTaq DNA polymerase (Applied Biosystems, CA). Final reaction concentrations were: 10 mM Tris-HCl (pH 8.3); 50 mM KCl; 2.0 mM MgCl2; 1 mg/ml BSA; 0.25 mM each of dATP, dCTP, dGTP, and dTTP; universal and species-specific primer concentration as shown in Table II; 0.2-4.0 ng template DNA (1/100 of DNA from entire adult mosquito)/reaction; and 1.25 U of Amplitaq/reaction. A Perkin Elmer Applied Biosystems 9700 thermocycler was used for PCR with the following parameters: initial denaturation at 95°C for 2 min followed by 35 cycles of denaturation at 95°C for 30 s, annealing at primer-specific temperatures shown in Table II for 30 s, and elongation at 72°C for 1 min, followed by a final extension at 72°C for 10 min. PCR product was separated on a 2.5% agarose gel and visualized with ethidium bromide stain. Fragment sizes were estimated by comparison to molecular weight standards provided by lambda DNA digested by Hind III, and phiX174 DNA digested with Hae III (Sigma, St. Louis, MO). RESULTS AND DISCUSSION Based on sequence derived from cloning of ITS2 (Li & Wilkerson, unpublished data, Table I) we designed a series of six ITS2 internal primers that can be used for identification of the four species in the complex (Table II). The six primers are used for single species identifications or, in the case of primers albCD and albABD, for groups of species. Single species-specific primers are run with both ITS2F and ITS2R primers. The combination of the ITS2F and ITS2R primers, along with a species-specific primer, gives a diagnostic band in the presence of target DNA template, but results only in an ~ 490 bp band, representing the entire ITS2 and flanking regions, if the target species DNA is absent. In all reactions, including those that give a species diagnostic band, the ~ 490 bp band can be expected. Note that it is intermittent when in combination with the species-specific primers as a result of competition for PCR components. Our primers were not tested on other An. (Nyssorhynchus) but, based on ITS2 sequence generated by us (unpublished data), and that in Genbank, no homologous annealing sites are apparent. The possibility of false positives is precluded however by the initial morphological identification. A reaction producing only the complete ITS2 band should result if a non-target species, either a member of the Albitarsis Complex or another Nyssorhynchus, was tested. Lack of any band indicates absence of or degraded DNA. Since sequence differences available for primer design were few we obtained spurious results if we used a multiplex of species-specific primers (data not shown). Because of this only one species-specific primer can be used per reaction. For the same reason, the reaction conditions presented here should be duplicated as closely as possible. The ABI 9700 PCR machine used by us was calibrated shortly before testing the primers, and we therefore believe that the temperatures reported here are accurate. Four primers allowed identification of single species and two serve to identify combinations of species (Table II). By following Fig. 1 it is possible to identify an unknown specimen that was first morphologically identified as belonging to the Albitarsis Complex. Primer albCD, with ITS2F and ITS2R, will either yield an ~ 390 bp fragment (often also with a ~ 490 bp fragment, see above) indicating the presence of either An. marajoara or An. deaneorum; or it yields only the ~ 490 bp fragment representing the entire ITS2 amplicon (Fig. 2), indicating the presence of An. albitarsis or An. albitarsis B. As can be seen in Fig. 1 an additional primer (always with both universal primers) is then chosen depending on the result: albC for An. marajoara (Fig. 3); or albB for An. albitarsis B (Fig. 4). The apparent negative results for An. de-aneorum and An. albitarsis are actually positive if judged by the first reaction, i.e. it has to be one or the other. If desired, a further confirmation reaction can be carried out for these two species; albD for An. deaneorum (Fig. 5), or albA for An. albitarsis (Fig. 6). In addition, primer albABD can be used to identify three species (Fig. 7), An. albitarsis, An. albitarsis B, and An. deaneorum. The two-step process may not be necessary if one can make a presumptive identification based on locality, e.g. only An. marajoara has been found north of the Amazon River, or morphology, head hair 3-C is multibranched only in An. deaneorum. Species-specific primers were tested with complete correlation on all specimens in our collection that had previously been identified using RAPDs, i.e. 99 An. albitarsis, 56 An. albitarsis B, 407 An. marajoara, and 41 An. deaneorum. An. marajoara from north of the Amazon River is of particular interest since Lehr et al. (2005) hypothesize the existence of an additional species ("E") in the Albitarsis Complex from Venezuela and the state of Roraima, Brazil. This finding was not corroborated by us with ITS2 sequence or through identifications using the technique described here. Recently, Merritt et al. (2005) also developed a method for identification of An. marajoara based on the presence of the fourth intron of the white gene, which is absent in the other members of the complex. We sequenced this fragment of white gene for a subsample of the specimens used here and agree with their results (data not shown). Our unpublished findings (Li & Wilkerson, see Table I Genbank accession numbers) show intragenomic variability in the ITS2 of these four species. Our primers were designed so that this variability would have no affect on PCR results. Note that because of indels and variable microsatellite regions PCR product size can vary, e.g. Fig. 7, An. deaneorum, "D" (see Genbank accessions in Table I). It is known that An. marajoara is a significant malaria vector, at least in Northeastern Brazil, but the status of the other species is poorly understood. The ITS2-based PCR protocol presented here provides a means for vector ecologists, malaria epidemiologists and control personnel to positively identify members of the Albitarsis Complex. ACKNOWLEDGEMENTS To the support of those who helped collect and rear many of the specimens used in this study. These include Terry Klein and Eric Milstrey, who were then members of the United States Army Medical Research Unit in Rio de Janeiro. But we especially thank Jose Bento Lima, without whose extra efforts many of samples would not have been available to us. To Lee Weigt at the Smithsonian Institution, Laboratory of Analytical Biology for use of laboratory and sequencing facilities and Jan Conn and Desmond Foley for thoughtful reviews of a draft manuscript. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05121t2.jpg] [oc05121t1.jpg] [oc05121f2.jpg] [oc05121f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}