 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 5, August 2005, pp. 501-513 Pre-mRNA trans-splicing: from kinetoplastids to mammals, an easy language for life diversity Mario Gustavo Mayer , Lucile Maria Floeter-Winter*/+ Laboratório
de Genética, Instituto Butantan, São Paulo, SP, Brasil *Departamento
de Fisiologia, Instituto de Biociências, Rua do Matão, travessa
14, 101, 05508-900 São Paulo, SP, Brasil Financial support: Fapesp, CNPq Received 7 April
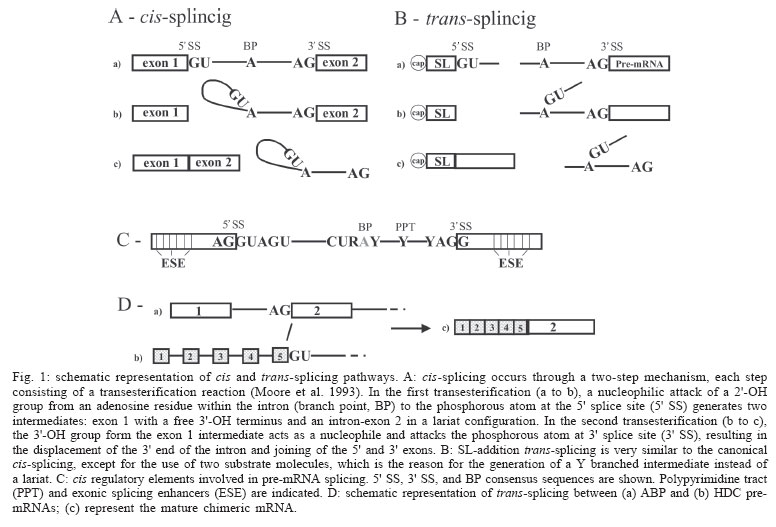
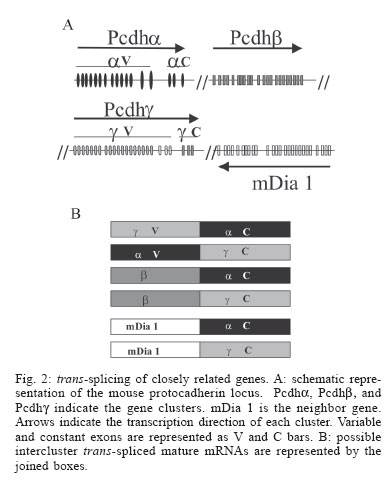
2005 Code Number: oc05122 Since the discovery that genes are split into intron and exons, the studies of the mechanisms involved in splicing pointed to presence of consensus signals in an attempt to generalize the process for all living cells. However, as discussed in the present review, splicing is a theme full of variations. The trans-splicing of pre-mRNAs, the joining of exons from distinct transcripts, is one of these variations with broad distribution in the phylogenetic tree. The biological meaning of this phenomenon is discussed encompassing reactions resembling a possible noise to mechanisms of gene expression regulation. All of them however, can contribute to the generation of life diversity. Key words: RNA processing - alternative trans-splicing - spliced leader RNA - trypanosomatids - nematodes - mammalian Most of the protein-coding eukaryotic genes display an interrupted structure alternating exons and introns. After transcription, introns must be removed from the primary transcript (pre-mRNA) to generate a translatable mature mRNA. It is interesting to note that the mature mRNA is constituted of 5' and 3' untranslatable regions (UTR) flanking the open reading frame (ORF) and that UTR are also exons. The precise excision of the introns and the joining of neighboring exons is a complex process generally named splicing, and when this processing occurs within a single pre-mRNA molecule it can also be called cis-splicing (Moore et al. 1993, Burge et al. 1999). Cis-splicing occurs in a two-step mechanism, each step consisting of a transesterification reaction (Moore et al. 1993) (Fig. 1A). The spliceosome, a ribonucleoprotein machinery composed of five small ribonucleoproteins (U1, U2, U4, U5, U6 snRNPs, named in relation to the kind of associated RNA molecule) and approximately 300 distinct proteins, is responsible for the splicing catalysis (Burge et al. 1999). Much effort was expended on the comprehension of the structure and dynamics of this complex machine during the splicing process, but the rules that discriminate introns and exons within the message are still poorly understood. Nevertheless, four conserved pre-mRNA sequence elements, which interact with the spliceosome, have been characterized as determinants in the splicing process (Moore et al. 1993, Burge et al. 1999) (Fig. 1C). In mammals, the 5' splice site consensus is AG/GURAGU (R for purine, /for splice site, bold for the dinucleotide 5' intron boundary) while the 3' splice site consensus is YAG/G (Y for pyrimidine, /for splice site, bold for the dinucleotides 3' intron boundary). There is also a conservation of nucleotide sequence around the branch point, CURAY (Y for pyrimidine, R for purine and A for branch point), and a polypyrimidine tract of variable length between the branch point and the 3' splice site. Recently, other cis-regulatory elements were implicated in splice site recognition, e.g. the exonic splicing enhancers (ESEs) (Fig. 1C). ESEs are localized within exons and interact with a family of proteins rich in serine and arginine (SR proteins) that recruits the spliceosome to the proximal splice site (Maniatis & Tasic 2002). Although the presence of those consensuses was observed, it is well known now that the splicing theme is full of variations. Yeast pre-mRNAs have a slightly different consensus for the 5' splice site (AG/GUAGU) than mammals, and the polypyrimidine tract is not as evident. Moreover, the discovery of other variations has broken some rules. Some introns, known as AU-AC, bear AU and AC at their 5' and 3' boundaries instead of the GU and AG dinucleotides, as well as different sequences around their branch point. Variations are not restricted to pre-mRNA sequences and involve the spliceosome machinery itself. A minor spliceosome presents in its composition U11, U12, U4atac and U6atac snRNPs instead of U1, U2, U4, and U6 snRNPs always found in major spliceosomes. The only snRNP that is common to both types of spliceosome is the U5 snRNP (Yu et al. 1999). The classical splicing mechanism involves the joining of exons encompassed in one pre-mRNA molecule (cis-splicing). One important variation of this picture can be found in early branched and at least in some pre-mRNAs of complex eukaryotes, where exons from distinct molecules could be joined together (trans-splicing) (Ullu et al. 1996). The following sections of the present communication intend to summarize the history, distribution, and possible biological meaning of the pre-mRNA trans-splicing process amongst different organisms. Trans-splicing: a brief history - In vitro studies using HeLa cell-free system showed for the first time that two independent transcripts could be joined together by trans-splicing (Konarska et al. 1985, Solnick 1985). Those data promptly suggested that the same trans reaction could take place in vivo in eukaryotic cells. The studies of antigenic variation in infective forms of the early-branched eukaryote Trypanosoma brucei, the causative agent of African trypanosomiasis, showed that all mRNAs encoding variant surface glycoproteins (VSGs) had an identical 39 nucleotide leader sequence at its 5' termini. That sequence, however, was absent in VSG genomic vicinities (Boothroyd & Cross 1982). Subsequent experiments identified the same sequence at the 5' termini in other mRNAs from T. brucei showing that the occurrence was not restricted to VSG genes, but should represent the addition of a leader sequence in all mRNA of that organism (De Lange et al. 1984). Further, similar sequences were found at 5' end of other trypanosomatids, showing that this phenomenon occurs in all those organisms (Parsons et al. 1984). Further research showed that the leader sequence is present in approximately 200 copies per genome, clustered in tandem, which do not map in the same chromosome as VSG genes (De Lange et al. 1983). Expression studies showed that transcription of the leader sequences generates a small RNA of 135-147 nt, which contains the 39 nt leader sequence at its 5' termini (Milhausen et al. 1984). Although these results exclude the occurrence of cis-splicing, other hypotheses were proposed to explain the structure of mature mRNAs of trypanosomes: (i) the pre-mRNA is transcribed using the leader sequence as initiator, and the resulting molecule is then processed by cis-splicing; (ii) the independently transcribed short RNA bearing the leader sequence is ligated, via its 3' end to the 5' end of the pre-mRNA and the resulting molecule is spliced in cis; (iii) the leader RNA and the pre-mRNA could be independently transcribed as in (ii) but the two substrates could be spliced in trans. Experimental analysis of pre-mRNA maturation detected a Y-branched intermediate molecule, instead of the typical lariat expected from cis-splicing, only compatible with a trans-splicing process (Fig. 1B) (Murphy et al. 1986, Sutton & Boothroyd 1986). The leader sequence was then named spliced leader (SL) or mini-exon, while the short transcript in the unprocessed form was named SL-RNA. The trans-splicing processing in trypanosomes is very similar to the canonical cis-splicing (Fig. 1A, B) (Ullu et al. 1996). The SL RNA is one of the substrates and presents the canonical GU splice junction at the 39 nt exon intron boundary. The other substrate is the pre-mRNA that contains the 3' canonical AG splice site localized in the non-translated 5' part of the molecule (Fig. 1B). This 5' region of the pre-mRNA also contains a conserved adenosine (A) residue that is used as the branch point in the process. Finally, a polypyrimidine tract is observed between the 3' acceptor site and the branch point. The discovery of natural trans-splicing in trypanosomes raised the question of how common this kind of processing could be among other organisms. In fact, just after those studies, trans-splicing by spliced leader addition was detected, for at least part of nematodes transcripts (Krause & Hirsh 1987). Nevertheless, it was not known if this kind of processing was part of the pre-mRNA processing repertoire of other eukaryotes. Computational search of the known features of trans-splicing substrates resulted negative (Dandekar & Sibbald 1990). Experiments in mammalian cells showed that SL sequences from nematodes and Leptomonas collosoma were accurately trans-spliced to both nematode and adenovirus suggesting that the machinery utilized in cis could perform a trans-splicing, therefore both machineries should present common features (Bruzik & Maniatis 1992). The natural occurrence of trans-splicing in mammalian cells was first time proposed for genes encoding immunoglobulin heavy chain, c-myb and androgen-binding protein (ABP) products (Shimizu et al. 1989, Sullivan et al. 1991, Vellard et al. 1991). For example, ABP is an androgen carrier protein produced in the testicular Sertoli cell but is also transiently expressed during rat liver development. The analysis of transcripts expressed in rat fetal liver showed existence of hybrid molecules between ABP and histidine decarboxylase (HDC) mRNAs (Fig. 1D). Then, it was proposed that trans-splicing generated the hybrid molecules, since the genes encoding these two proteins are localized in different chromosomes. Moreover, the joining sequence between the two mRNAs occurred at nucleotides preceding a canonical donor site (/GU) at the fifth ABP intron and following acceptor (AG/) site of the HDC intron 1 (Fig. 1D). Although these experiments did not address the mechanisms whereby the mammalian hybrid pre-mRNA arises, it suggests that trans-splicing could be the responsible one. At the present time, the pre-mRNA trans-splicing in eukaryotic cells is more often detected, suggesting its participation in distinct biological processes. It is important to highlight that in early branched eukaryotes trans-splicing is responsible for the joining of a leader sequence that is not translatable, while in mammalian cells trans-splicing could generate a different protein product, enhancing protein diversity in the cell. Thus, it is expected that these differences might have different biological meanings. Trans-splicing of viral transcripts: cryptic sites impacting cell viability? - The analysis of the transformation potential of different SV40 fragments in rat cells provided the first evidences that mammalian cells were able to join two independently transcribed viral pre-mRNA molecules by trans-splicing. SV40 genome is divided into early and late regions, the expression of the early region is required for the induction and maintenance of the transformed cell state. This region encodes the large T and the small t antigens, which are generated by alternative "cis-splicing". Two different 5' donor sites and one 3' acceptor site are used for the production of the two mature mRNAs. Microinjection experiments in rat cells using a plasmid construct bearing the distal part of the large T intron, the small t intron and the second large T exon under the control of the early SV40 promoter resulted in the production of two mature mRNAs (Eul et al. 1995). One of them is the expected mRNA (T2 mRNA), processed by cis-splicing with the excision of the small t intron. The other transcript detected (T1 mRNA) is the result of the use of the canonical cryptic 5' donor splice site, located in the second large T exon to join the small t exon using its 3' acceptor site. Since the 3' acceptor site precedes the 5' donor site, the only possibility for the generation of T1 transcripts is a trans-splicing reaction. Interestingly, T1 and T2 mRNAs could be translated producing two distinct antigens, which in turn suggests a possible biological function for the process. Subsequently the spectrum of virus transcripts that could be processed by trans-splicing experiments was broadened, now indicating the joining of two distinct molecules. Mammalian cells microinjected with in vitro produced pre-mRNAs from HIV-nef gene and SV40 T antigen were able to produce hybrid mRNAs (HIV-nef/T-antigen) as well as fusion proteins (Caudevilla et al. 2001b). Both 5' cryptic splice sites of the HIV-nef mRNA were spliced to the 3' acceptor site common to the large T and small t antigens. Moreover, for the first time, it was possible to detect hybrid molecules between viral (HIV-nef mRNA) and cellular transcripts, generated by trans-splicing. Eight different HIV-nef/cellular hybrid mRNAs were detected. In four of those products, the HIV-nef transcript contributed with one of their cryptic donor sites while cellular transcripts contributed with the acceptor site (Caudevilla et al. 2001b). Although the above data demonstrate that mammalian cells can utilize viral transcripts as trans-splicing substrates in generating hybrid protein, the biological significance of these products was not addressed as well as its occurrence during the course of an infection. However, the impact of viral infection on host RNA splicing could be evaluated by the detection of a great variety of cellular transcripts, including glyceraldehyde-3-phosphate dehydrogenase (GADH) and b-actin, and exon 2 of the major late transcript of adenovirus (MLT), in the course of the infection (Kikumori et al. 2002). A construction containing the 3' acceptor site of MLT intron 1, a c-myc epitope, a polyhistidine tag, and a polyadenylation signal of bovine growth hormone was transfected in mammalian cells. Trans-splicing of the construct transcript with cellular RNAs could generate tagged proteins with a third of possibilities because of the start codon frame. The tagged proteins were in fact detected by immunoprecipitation in higher levels than controls, but no apparent specificity was observed, suggesting that trans-splicing mediated by the 3' acceptor site of MLT intron 1 could be unregulated. Although these experiments failed to find a specific protein related to cell viability, the use of a tetracycline-inducible system showed that this promiscuous trans-splicing has a discrete but significant impact on cell growth (Kikumori et al. 2002). In the examples mentioned above, the cryptic sites constitute intrinsic characteristics of the genes that lead them to participate in trans-splicing process, and are not associated with point mutations of the regular splice sites (Caudevilla et al. 2001b). Since the number of known viral cryptic splice sites is still low, it is unclear if there are other attributes which direct these genes to engage in trans-splicing mechanisms. While the steps of heterologous trans-splicing reactions during viral infection are well established, its biological meaning is just being defined. In fact, the trans-splicing could represent a transgression in cellular RNA processing affecting the cell growth, and a consequent mechanism for adenoviral-mediated cell death (Kikumori et al. 2002). It is not clear if specific cellular mRNAs could act as targets of trans-splicing producing hybrid proteins involved in cell growth and death (Caudevilla et al. 2001b), or if the effect in cell viability may be non-specific, and related to the interference in the overall regular cis-splicing in the cell (Kikumori et al. 2002). Mammalian interchromosomal trans-splicing: absence of consensus splicing sites - Genes localized at distinct chromosomes can be the templates for the transcription of two pre-mRNAs engaged in the interchromosomal trans-splicing, and as described above for the ABP gene (Sullivan et al. 1991), other examples of mammalian interchromosomal trans-splicing have been proposed. Hybrid mRNAs have been detected among transcripts of the genes encoding human calcium/calmodulin dependent protein kinase II and signal recognition particle 72, respectively on chromosomes 10q22 and 18 (Breen & Ashcroft 1997); human acyl-Coa:cholesterol acyltrans-ferase-1 (ACAT-1) and Xa exon, respectively on chromosomes 1 and 7 (Li et al. 1999); rat leukocyte common antigen-related (LAR) tyrosine phosphatase receptor and a 3' UTR on chromosomes 5 and 1 respectively (Zhang et al. 2003), and the mouse meiotic recombination gene Msh4 localized on chromosome 3 and three different sequences on chromosomes 16, 2, and 10 (Hirano & Noda 2004). It is noteworthy that GU-AG rule is not obeyed most of the proposed interchromosomal trans-splicing. In fact, the chimeric ABP-HDC transcript is the unique example in which the GU-AG rule could be used for the proposition of a conventional trans-splicing processing. For example, the 5' and 3' intron boundaries proposed for the generation of b, d and e variants of the Msh4 gene are respectively UG-GU, AU-AU, and UC-CA (Hirano & Noda 2004). However, it is interesting to note that the d splice donor and acceptor sites are similar to the AU-AC introns. The almost absence of conventional splicing boundaries in the generation of hybrid mRNA molecules coded by genes located in different chromosomes suggests an alternative mechanism of RNA processing for their generation. However, trans-splicing cannot be completely excluded if it is assumed that it occurs through a non-conventional splicing mechanism. So, the study of mechanisms involved in the generation of such hybrid molecules is important for the distinction between a splicing related and other type of RNA processing. Hybrid mRNAs transcribed from genes in different chromosomes have also been detected in chicken and rice seeds (Vellard et al. 1991, Kawasaki et al. 1999). In chicken, hybrid mRNAs are formed between the transcript of c-myb proto-oncogene localized on chromosome 3 and an exon on chromosome 17 (Vellard et al. 1991). In rice, calcium dependent seed-specific protein kinase mRNA is coded by two regions located on chromosomes 6 and 10 (Kawasaki et al. 1999). In humans, we will highlight the case of ACAT-1. This is a relevant protein in lipid metabolism. Two isoforms of the protein were detected in in vitro expression studies (Yang et al. 2004). One of those isoforms is encoded by a trans-spliced mRNA, in which the initiation codon is GGC (Gly) instead of AUG (Met), suggesting a possible biological function related to its diversity. However, ACAT-1 mRNA is rare and the protein produced from the trans-spliced RNA could be detected only in macrophages stimulated with phorbol esters (PMA) and in human monocyte-derived macrophages. Moreover, the activity of the trans-spliced produced protein is approximately 30% of the other isoform. As the abundant isoform forms tetramers, the possible formation of heterotetramers could constitute a negative regulatory mechanism. Intergenic trans-splicing of closely linked genes in mammals: lower frequency or a noise reaction? - Another possibility in the generation of hybrid mRNA molecules is trans-splicing of transcripts coded by a cluster of genes. Hybrid mRNAs were detected between members of the immunoglobulin locus (Shimizu et al. 1989, Fujieda et al. 1996), the human GTPase RSG12 gene and a sequence localized 170 kb downstream from the RGS12 gene (Chatterjee & Fisher 2000), transcripts coded by genes of the cytochrome P450 3A cluster (Finta & Zaphiropoulos 2002), and members of the mouse proto-cadherin locus (Tasic et al. 2002). The four cytochrome P450 3A genes (CYP3A4, CYP3A5, CYP3A7, and CYPA43) are located in a cluster of human chromosome 7 (Finta & Zaphiropoulos 2002). CYPA43 is in a head-to-head orientation to the CYP3A4 and CYP3A5 genes, i.e. they are transcribed from different DNA strands. RT-PCR and RNase protection studies showed hybrid molecules between the first exon of CYPA43 and CYP3A4 or CYP3A5 exons. The joining of intergenic exons occurs at canonical splice sites and generates translatable mRNAs, however endogenous protein products were not detected. Moreover, quantitative RT-PCR experiments showed that the expression levels of hybrid molecules is 650:1 in relation to the canonical the mRNAs. Mammalian protocadherins (Pcdh) are a family of cell surface proteins that could enhance neuronal protein diversity. Pcdh gene families (a, b, and g) are clustered in the genome. The three clusters are localized in a 900 kb region of mouse chromosome 18 (Fig. 2A). Mouse Pcdh a contains 14 variable exons while Pcdh g contains 22 variable exons. Each variable (V) exon encodes chaderin extracellular domais, the transmembrane portion and a small piece of the cytoplasmic domain. The three constant exons are located at the 3' end of each cluster and encode the C-terminal part of the cytoplasmic domain. Pcdh pre-mRNAs are processed by splicing of one V exon to the first constant exon of each cluster. Otherwise, the mouse Pcdh b cluster contains 22 V exons and do not have a constant exon that in theory could encode a cytoplasmic domain, i.e. each single exon encodes a Pcdh b protein. It was shown that each a and g V exon is transcribed through its own promoter, and the promoter choice determines the exon which will be spliced to the first constant exon (Tasic et al. 2002). The a and g isoforms of Pcdh are generated by alternative cis-splicing within a gene cluster, and interchromosomal and intracluster trans-splicing was excluded (Tasic et al. 2002). However, intercluster trans-splicing could be detected between a or g V exons and the first constant exon of the g or a respectively, as well as b exons and exons from a nearby gene mDia1 can trans-splice to the first constant exon of a or g clusters (Tasic et al. 2002) (Fig. 2B). This finding suggests the possibility of increase in diversity of Pcdh proteins by intercluster trans-splicing, although the levels of trans -spliced mRNAs are two orders of magnitude lower than the cis-spliced mRNAs, and then functional relevance has to be demonstrated. The correlation between promoter activity and the choice of the exon that will be the first in the mature mRNA could be interpreted as an interaction between transcription and splicing (the coupling model) with the recruitment of the splicing machinery to the capped proximal exon (Tasic et al. 2002). The analysis of protocadherin transcripts showed that only the capped-proximal V exons could be cis-spliced or trans-spliced to the first constant exon (Tasic et al. 2002). For a trans-spliced product, the coupling model cannot be applied since in trans-splicing reactions, both pre-mRNAs are independently transcribed. In trans-splicing reactions the 5' splice site must pair with 3' splice site from another molecule, and according to the coupling model it would be possible that capped-proximal 5' splice site transiently dissociates from RNA polymerase CTD domain before the synthesis of the 3' splice site, rendering 5' splice site available for paring with another free 3' splice site. In this case, coupling transcription and splicing could minimize inappropriate trans-splicing, explaining the low levels of trans-spliced mRNAs, and suggesting that trans-splicing could be a noise in cis-splicing reactions. This proposal is mainly supported by two facts: the low frequency of trans-splicing products and the absence of a biological function assigned for them. Mammalian trans-splicing and exon repetition: efficient but non-essential - Exon repetition is a term that describes the presence of more than one copy of an exon in mRNAs without alterations at the DNA level. It was proposed that trans-splicing between two identical pre-mRNA molecules could be involved since the joining of duplicated exons are precise. Exon repetition was first described during expression studies of two rat genes, carnitine octanoyl transferase (COT) and medium chain acyl-CoA synthetase (SA) (Caudevilla et al. 1998, Frantz et al. 1999). After that, the observation was made in a small number of other rat and human genes including the rat sensory neuron specific (SNS) voltage-gated sodium channel (Akopian et al. 1999), the human and rat Sp1 transcription factor (Takahara et al. 2000, 2002) and the hERa human estrogen receptor-a (Flouriot et al. 2002) genes. The COT gene is composed of 17 exons. During studies of COT gene expression in Sprague-Dawley rat liver, two cDNAs with exon duplications were obtained, i.e. instead of the canonical organization exon1-exon2-exon3…exon17, these cDNAs presented the alternative structures exon1-exon2-exon2-exon3…exon17 or exon1-exon2-exon3-exon2-exon3…exon17 (Akopian et al. 1999). An in silico analysis of putative protein synthesized from the canonical cis-spliced form showed a 70 kDa protein, but in relation to the exon repeated transcripts, only the longest trans-spliced alternative form (exon2-exon3-exon2-exon3) is in frame to be translated in a larger (80 kDa) protein. In fact, COT specific antibodies detected two proteins with apparent molecular mass of 69 and 79 kDa, in Western blot experiments for peroxisomal proteins, indicating a putative protein product translated from a possible intragenic trans-spliced mRNA. Remarkably, the analysis of COT gene exon repetition in mammalian related species showed it is not conserved, demonstrating that it is not essential, at least for the studied species. The absence of exon repetition in these mammals could be explained by the loss of an ESE sequence in the exon 2, which is present in the rat (Caudevilla et al. 2001a). A more complex study of exon repetition using two rat lines showed that Sa is a gene that is expressed in the liver and kidneys of rats. The transcript is more abundant in spontaneously hypersensitive rats (SHR) than the normotensive Wistar-Kyoto rats (WKY), although exon repetition was only detected in WKY rats. The observed pattern was exon1-exon2-exon2-exon3-exon4-exon5-exon6 and exon1-exon2-exon3-exon4-exon2-exon3-exon4-exon5-exon6 (Frantz et al. 1999). Remarkably, relative frequency of exon repeated transcripts is as abundant as the canonical ones showing an efficient production of the trans-spliced molecule (Rigatti et al. 2004). The concomitant analysis of COT exon 2 nucleotide sequences from these two rat lines showed that both sequences were identical. Moreover, as for the Sa transcripts, COT exon 2 repetition was observed only in WKY rats, raising the possibility that trans-splicing could be determined by a trans-acting factor and not by a ESE. However, segregation studies using SHR and WKY rats showed that exon repetition is restricted to specific alleles (Rigatti et al. 2004), i.e. the determinant factor for COT transcript exon repetition is a cis-regulatory element, but not related to the proposed ESE sequence. A possible biological role for exon repetition is related to proteome diversity. The only putative product detected is the 80 kDa protein coded by exon1-exon2-exon3-exon2-exon3-exon4-exon5-exon6 COT mRNA. However, this protein is not essential, since exon repetition was not observed in all rat lines studied (Rigatti et al. 2004). Additionally, the COT transcript bearing only the exon 2 repetition is out of frame. It was also observed that a construct bearing exon 2 repetition dropped the expression of the reporter gene, a fact that was interpreted by the authors as a detrimental effect of the exon repetition in natural COT expression (Rigatti et al. 2004). On the other hand, a negative regulation could also be proposed to explain the biological meaning of this exon repetition. In the absence of other studies related to the detection and function of protein products coded from transcripts bearing repeated exons, it could be argued that the generation of these transcripts could represent a noise reaction. On the contrary, a hallmark of the transcripts bearing repeated exons, besides the low number of genes studied, is their high level of expression (Caudevilla et al. 1998, Rigatti et al. 2004), suggesting a limited but efficient process. Trans-splicing in Drosophila: efficient and essential - In contrast to the non-essential trans-splicing described for mammals, studies of two Drosophila loci showed that the phenomenon generate diversity in the production of two essential proteins (Dorn et al. 2001, Labrador et al. 2001, Horiuchi et al. 2003). The Drosophila modifier of mdg4 [mod(mdg4)] locus encodes a large number of proteins with different functions especially related with the formation of chromatin complexes (Dorn et al. 2001, Labrador et al. 2001). The observed diversity could be explained by a combination of common first four exons and 26 different terminal exons. Upstream sequences of the fifth exons present AG dinucleotides at the 3' introns boundary and the putative branch point and polypyrimidine tract. As seven out of 26 different 3' ends were encoded on the opposite strand, it was proposed that trans-splicing was responsible for the mRNA diversity (Dorn et al. 2001). Fifth 3' exon originated from each opposite or same strand were inserted in a different chromosome of the endogenous locus, to construct two distinct transgenic flies. RNA processing analysis of those flies confirmed that both types of exons were joined to the first four exons, suggesting that trans-splicing could account for the generation of all mod(mdg4) isoforms. Similarly, when the first four exons bearing its own promoter were expressed on a distinct chromosome, a mutation in the corresponding endogenous sequence could be rescued. All observation in this artificial system showed that chromosomal context is not important for the generation mod(mdg4) mature mRNAs. Moreover, multiple TATA-box-containing elements were found throughout the entire locus, and at least one of the fifth exons that were transcribed from the same strand of the common exons had its own promoter function determined. The results suggested the existence of independent transcriptional units for each fifth exon, although the other 25 promoters were not characterized. The Drosophila longitudinals lacking (lola) complex gene (Horiuchi et al. 2003) encodes at least 20 isoforms of BTB-Zn finger transcription factors required for axon guidance decisions in the Drosophila nervous system development. The protein is translated from a mRNA variant that contains a constant region (C) composed of exons 5-8, which encodes a N-terminal BTB dimerization domain followed by one or two variable exons (V), which encodes the C-terminal zinc finger variable domain. To complicate the scenario, exons 1 to 4 are alternatively used as the initial 5' exon through an alternative transcription initiation from four possible sites. So, the complex locus spanning 60 kb, where 32 exons are aligned on the same DNA strand, can generate 80 splicing variants by both alternative transcription initiation and alternative splicing. Alternative splicing of the following variable exons (9-32) produces 20 different combinations, 17 of them have unique zinc finger motifs, suggesting different target DNA sequences. The isoforms have a complex pattern of expression, and expression of different isoforms in the same cell is observed, multiplying functional diversity once dimerization of different isoforms could occur. Mutations where a specific lola isoform was inactivated correlate to a specific defect in axon guidance choice point, suggesting that alternative splicing in lola locus is important in the determination of axon trajectories. Lethal mutations in constant exons could complement mutations in variable exons localized in different homologous chromosomes. RT-PCR experiments showed that wild type chimeric mRNAs were detected in F1 flies heterozygotes for mutations in V and C exons. Moreover, the joining between the V and C exons synthesized by the two homologous chromosomes occurred at canonical splice-sites (Horiuchi et al. 2003). It is noteworthy that approximately half of the mature mRNA for some isoforms is originated by trans-splicing between pre-mRNAs transcribed from two homologous chromosomes. Also, it was found that at least one of the variable trans-spliced exons is probably transcribed from its own promoter. A negative position effect for the frequency of trans-splicing was observed in flies bearing an inverted locus with respect to its homologous. This observation, more pronounced if the locus was positioned in a distinct chromosome, suggests that chromosome pairing during transcription, a typical feature in insects, is important for the generation of trans-spliced lola transcripts. The position interference observed in trans-splicing of the lola transcripts is not observed in modifier of mdg4 [mod(mdg4)] locus, since trans-splicing of modifier of mdg4 [mod(mdg4)] pre-mRNAs occurred when the transcripts were expressed from different chromosomes (Dorn et al. 2001). However, the expression system of the transgene, GAL4-UAS system, does not represent the natural situation, and different levels of transgene transcript could be produced which escape a natural regulation (Horiuchi et al. 2003). SL trans-splicing: a constitutive reaction - The classical trans-splicing consists in the addition of a short SL sequence to the 5' UTR of pre-mRNAs. Although described at first in Kinetoplastida, SL trans-splicing was subsequently shown to occur in other protists of the Euglenozoa phylum, i.e. Euglenida (Tessier et al. 1991, Ebel et al. 1999, Frantz et al. 2000), Diplonemida (Sturm et al. 2001), and Kinetoplastida (Murphy et al. 1986, Sutton & Boothroyd 1986, Laird et al. 1987). In the metazoans, trans-splicing has been described in free-living or parasitic nematodes (Krause & Hirsh 1987, Blaxter & Liu 1996), in trematodes (Rajkovic et al. 1990, Davis et al. 1994) and cestodes (Brehm et al. 2000) as well as in turbelarians (Davis 1997) of the Platyhelminthes phylum. More recently, this form of RNA processing has been described in Hydra, a member of Cnidaria phylum (Stover & Steele 2001) and, surprisingly in two members of the Urochordata subphylum of chordates, the ascidian Ciona intestinalis (Vandenberghe et al. 2001) and the appendiculariam Oikopleura dioica (Ganot et al. 2004). It is absent in a variety of organisms where EST libraries were intensively sequenced, e.g. arthropods, plants, most protists, fungi, and vertebrates. This wide but sporadic range of phylogenetic distribution among eukaryotes prompted the question if SL trans-splicing arose independently many times or if it was originated once and lost from various lineages (Nilsen 2001, Blumenthal 2004). At present, based on phylogenetic data it is impossible to discriminate between both hypotheses. It is important to stress here that determining a representative phylogenetic distribution of SL trans-splicing in eukaryotes is not an easy task because there is little sequence conservation among the identified SL RNAs from the various phyla. A directed EST project intended to cover a wide phylogenetic range could in theory help in the determination of SL trans-splicing distribution (Nilsen 2001). The poor sequence conservation in SL RNAs across the diverse phyla is accompanied by a little length conservation, although they are invariably small RNAs (< 150 nt) (Nilsen 2001). The mini-exon sequence, which is transferred to the 5' end of pre-mRNAs, is also variable in length and ranges from 16 nt to 51 nt (Davis 1997, Vandenberghe et al. 2001). Most of the SL RNAs showed a variable secondary structure. Nevertheless, a common secondary structure composed of three stem-loops is observed in kinetoplastids and nematodes (Nilsen 2001). SL trans-splicing and cis-splicing are very similar and little differences were observed to date. Intron boundaries are defined by the same elements observed in cis-splicing, although the 5' donor splice site is localized in the SL RNA molecule and the 3' acceptor splice site is localized in the pre-mRNAs. The 5' intron boundary is defined by a GU dinucleotide, while the 3' intron boundary is defined by an AG dinucleotide (see Fig. 1). A polypyrimide tract is observed upstream of the AG dinucleotides in kinetoplastids (Liang et al. 2003), cnidarians and in C. intestinalis (Vandenberghe et al. 2001), whereas in nematodes there is no polypyrimidine tract associated to the 3' splice site. In nematodes a conserved sequence UUUCAG/ (AG, acceptor site) is required for proper processing (Conrad et al. 1993, Romfo et al. 2001). Finally, in both processes it is possible to find out the participation of ESEs, SR proteins as well as similar spliceosome which are formed by almost the same snRNPs and proteins (Sanford & Bruzik 1999, Liang et al. 2003). The difference between the two types of processing resides in the components of the 5' splice site recognition. While U1 snRNP is required in cis-splicing, it does not participate in SL trans-splicing. In fact, the SL RNA acts as a substrate as well as a component of the spliceosome catalytic complex (Liang et al. 2003). Moreover, in nematodes the SL snRNP contains two proteins that are exclusive to the SL snRNP of nematodes and essential to the SL trans-splicing but not for cis-splicing (Denker et al. 2002). In kinetoplastids SL trans-splicing processes virtually all pre-mRNAs, while, only one gene was found to have its mRNA processed by cis-splicing (Mair et al. 2000). This prevalence raised the question of the biological role of trans-splicing in those organisms. Kinetoplastids present polycistronic transcription. Moreover, a transcriptional analysis of two Leishmania major chromosomes showed few promoter regions to drive the expression of their protein coding genes (Martinez-Calvillo et al. 2003, Worthey et al. 2003). In L. major chromosome 1 the transcription starts at a single strand-switch region and goes bi-directionally towards each telomere (Martinez-Calvillo et al. 2003). An analogous analysis of chromosome 3 showed that transcription starts near each telomere towards a region occupied by a tRNA gene at which transcription terminates (Worthey et al. 2003). These observations clearly demonstrate that the transcription units in kinetoplastids are very long and SL trans-splicing functions in the individualization of the messages. A single type of SL RNA is trans-spliced to every cistron with the peculiarity that polyadenylation is coupled to the splicing process (LeBowitz et al. 1993). In fact, in these organisms a polyadenylation cleavage site is not a consensus but it is determined by a fixed distance in relation to the downstream polypyrimidine tract of neighbor cistron (Matthews et al. 1994). The trans-splicing of the downstream gene occurs before the polyadenylation of the upstream sequence (LeBowitz et al. 1993). Part of nematode C. elegans genome is transcribed as polycistronic units and can be related to the prokaryote operons (Blumenthal & Gleason 2003). In prokaryotes polycistronic transcription results in a single multi-gene mRNA, which is translated into distinct proteins on ribosomes. The operon structure allows the co-regulation of genes of the same metabolic pathway. In C. elegans, the polycistronic transcript is first processed by trans-splicing and then transported to the cytoplasm to be translated. C. elegans has two types of SL RNAs involved in trans-splicing, namely SL1 and SL2. SL1 is responsible for the processing of the majority of pre-mRNAs engaged in trans-splicing, mostly those not organized in operons. For the pre-mRNAs organized in operons, SL1 is trans-spliced to the 5' end of the first cistron, i.e. it is trans-spliced to the acceptor site nearest to the promoter. SL2 is then used in trans-splicing the following downstream regions, i.e. it is a specialized form of SL whose function is to generate individual mRNAs from a polycistronic precursor. It was demonstrated that approximately 15% of C. elegans genes are organized in operons and more than 90% of these genes are SL2 trans-spliced (Zorio et al. 1994, Blumenthal et al. 2002). In fact, there is another type of operon, present in relatively small number, in which SL1 is the only form of SL RNA utilized to process the polycistronic transcript (Blumenthal 2004). The individualization of mature mRNAs involving SL trans-splicing in C. elegans is very similar to the kineto-plastid processing, although differences exist. In C. elegans operons the first step is the polyadenylation of the upstream sequence to which follows the trans-splicing of the downstream transcript. The AAUAAA poly-adenylation signal is approximately 100 bp apart from the downstream acceptor site, and its destruction decreases dramatically the trans-splicing of the downstream gene (Blumenthal 2004). Operons were found in the trematode Schistosoma mansoni (Davis & Hodgson 1997) and recently, in the urochordate O. dioica (Ganot et al. 2004). The operon organization, likewise for C. elegans, is related to SL trans-splicing. These data suggest that in eukaryotes one strategy for individualizing mRNAs from long polycistronic transcripts is SL trans-splicing. Another important feature of SL RNAs is that they have a 5' cap structure. The addition of m7G is essential for eukaryotic RNA metabolism and processing. In the nucleus, it is involved in pre-mRNA splicing and in directing mRNAs and UsnRNAs to the cytoplasm. In the cytoplasm, it is related to mRNA stability and translational initiation. So, the transfer of SL sequence to the 5' UTR of pre-mRNAs results in the capping of the mRNA, which persists during the mature mRNA life (Tschudi & Ullu 2002). Thus a SL trans-splicing reaction can be considered as a trans-capping reaction, making part of the repertoire of biological roles of SL trans-splicing. In kinetoplastids, the m7G is linked by a 5'-5' triphosphate bridge to the first SL sequence nucleotide (Tschudi & Ullu 2002). The cap structure however is more complex and is named cap 4 because the first four 5' nucleotides adjacent to the cap are modified by methylations. By analogy with other eukaryotic biological systems, the cap 4 structure was always implicated in mRNA stability, transport and translational regulation. A few indirect evidences support this analogy, e.g. mutant pre-mRNAs that do not trans-splice efficiently do not accumulate in vivo (Ullu et al. 1996). A mutational analysis of SL sequence showed that the part of the sequence itself and/or cap 4 formation are relevant for the association of mature mRNAs with the polysomes, suggesting the importance of this structure in the translation of kinetoplastid pre-mRNAs (Zeiner et al. 2003). Finally, in vitro analysis of cap 4 binding to recombinant eIF4E translation factor of Leishmania demonstrates its association with the translation machinery (Lewdorowicz et al. 2004). In nematodes SL 1 function is not associated to the individualization of polycistronic transcripts. In C. elegans the predominant form of trans-splicing is SL1 trans-splicing (57%) (Zorio et al. 1994) as well as for Ascaris suum (70%) (Maroney et al. 1995). In Hydra (Stover & Steele 2001) and C. intestinalis (Vandenberghe et al. 2001) there is no described operon structure to date. Taken altogether, these observations suggested that another role for SL trans-splicing exists in these organisms. SL1 and SL2 RNAs of C. elegans, and the SL RNAs of Hydra and C. intestinalis have a common cap structure, the N-2,2,7 trymethylguanosine cap (m32,2,7GppG or TMG cap). As it was suggested for kinetoplastids, it was postulated that SL structure and TMG cap could modulate the translational efficiency of mRNAs. Recently, the effect of trans-splicing on translation efficiency was assayed in Ascaris embryo cell-free translation system showing that each TMG cap or spliced leader sequence alone decreased the activity of a reporter gene (Lall et al. 2004). However, both features act synergistically to promote efficient translation, suggesting that SL trans-splicing has a biological role in promoting translation. So, resolving polycistronic transcripts and promoting translation are two biological functions recognized up to now for SL trans-splicing. A more precise map of the phylogenetic distribution of trans-splicing, as well as the functional analysis of this type of processing will probably solve the questions of its origin and other putative function(s). Alternative SL trans-splicing: variability or noise? - The majority of SL trans-splicing events so far described in kinetoplastids pointed to a precise use of 3' acceptor sites. However, many trials have shown that more than one acceptor site within the same intergenic region can be used in the generation of mature mRNAS coding the same ORF but distinct 5' UTR extensions (Vassella et al. 1994, Nepomuceno-Silva et al. 2001, da Silva et al. 2002, Manning-Cela et al. 2002). Expression studies of the single copy arginase gene in the kinetoplastid L. (L.) amazonensis showed that SL was added to more than two different acceptor sites in the pre-mRNA (da Silva et al. 2002) (Fig. 3A). Nucleotide sequence analysis of the longest cDNA demonstrated that two consecutive AG dinucleotides (positions 272 and 270) can be used as acceptor sites, while sequencing of the short cDNA showed that another two acceptor sites (positions 137 and 129) can either be used (da Silva et al. 2002). Thus the two far apart regions can be used as acceptor sites, each one displaying a local micro-heterogeneity. Interestingly, each one of the two acceptor regions is preceded by a polypyrimidine tract, an important element for 3' splice acceptor site determination. The role for the presence of more than one 3' acceptor site in the arginase gene is uncertain, however it could be related to an evolutionary drive that permits the accumulation of acceptor sites upstream of the ORF, which guarantee the pre-mRNA processing by trans-splicing. In kinetoplastids, alterations in acceptor site usage of one gene could implicate in alternative polyadenylation sites of the upstream gene. Since there is no polyadenylation signal sequence and polyadenylation of the upstream gene occurs at a ~100 nt distance subsequently to the trans-splicing of the downstream transcript. The alternative acceptor sites could be used as a regulatory mechanism for the expression of upstream genes. It is important to highlight here that gene expression in trypanosomatids are mainly post-transcriptionally regulated, and different 5' UTR and 3' UTR extensions could contribute with this regulation by generating different UTR structures and/or different targets for the binding of factors. Alternative acceptor splice sites were also found for Trypanosoma cruzi TcRho1 and Lyt1 transcripts (Nepo-muceno-Silva et al. 2001). TcRho 1 is a GTPase member of the Ras superfamily, which are involved in diverse signal transduction pathways. The analysis of 5' UTR of TcRho1 transcripts showed that five alternative AG splice acceptor sites are used to generate five different mature mRNAs. Two acceptor sites were mapped in the intergenic region but the other three were found within the ORF. The alternative ORFs downstream of the three TcRho1 ORF internal acceptor sites could direct the synthesis of putative small protein 54 residues long. Nevertheless, no similar proteins were detected in protein databanks. Two different polypyrimidine tracts were detected, one in a region localized upstream of the intergenic acceptor sites while a short polypyrimidine tract is present upstream of the three ORF internal acceptor sites. As proposed for the Leishmania arginase gene, a similar biological role could be attributed for the intergenic acceptor sites in T. cruzi TcRho1 gene. On the other hand the utilization of acceptor sites inside ORFs could contribute with a negative post-transcriptional regulation of gene expression since it would generate truncated mRNA (Nepomuceno-Silva et al. 2001). An alternative explanation for the biological meaning of acceptor sites inside ORFs emerged from studies of the LYT1 transcripts in T. cruzi. LYT1 protein (LYT1p) participates in diverse, apparently non-related, biological processes (Manning-Cela et al. 2002). Alternative trans-splicing produced three different transcripts; two of them were processed in the intergenic region while a third type was processed inside the ORF (Fig. 3B). One of the transcripts processed in the intergenic region is present in lower amounts than the others and its acceptor site is a non-canonical GG dinucleotides. The ORF analysis showed a putative signal sequence at its N-terminal part and a nuclear localization sequence at its C-terminal moieties. The alternative addressing of LYT1p to the nucleus or to the cell surface could explain the diverse roles attributed to this protein. It is interesting to note that the transcript processed inside the ORF would be translated in a truncated protein that lacks the signal sequence. Moreover, the amount of the two major transcripts is regulated through the parasite development. This way the alternative trans-splicing generates two transcripts that could be translated in two different proteins with apparent different localization in the cell and participating in different processes (Manning-Cela et al. 2002). Alternative SL-trans-splicing in kinetoplastids is a mechanism able to produce different transcripts from the same pre-mRNA. Otherwise, the utilization of non-canonical acceptor sites could simply reflect a noise in the major reaction. A different splice acceptor site was mapped in the intergenic region of the proton translocating P-type adenosine triphosphatase LDH1A of L. donovani and was pointed to be a result of an allelic variation, but it only occurred when the cells were maintained for a high number of culture passages (Fig. 3C). So, care should be taken in relation to allelic variations and/or variant appearance in long term culture stocks (Stiles et al. 1999). Interestingly, these results correlate with the observation, in T. cruzi, of a mutation that resulted in the loss of the canonical AG and lead to the use of the next AG dinucleotide as an acceptor (Hummel et al. 2000). It is interesting to note that cis and trans-splicing occur in the same nuclear environment. A tight regulation was shown in C. elegans where cis-splice donor sites are dominant over SL-donor site. However, trans-splicing at cis-acceptor sites have been described in trypanosomes, trematodes and cestodes, as recently discussed by Hastings (2005). Those observations could represent another way of generating transcript variability. As a conclusion, splicing is an important regulatory mechanism in the development of organisms and also a key to assure genetic variability. Post genomic information has been showing that the number of expected genes in many organisms is lower than the estimated. Part of the variability is generated at the level of RNA molecules. Alternative trans-splicing of transcripts could contribute to the expansion of protein diversity by raising the possibilities of exon combinations. In mammals, the features considered to engage two independently transcribed pre-mRNA molecules in trans-splicing are the absence of DNA recombination and the joining of RNA sequences through the use of putative canonical splice sites. Interchromosomal trans-splicing is the most difficult processing to demonstrate, since just one of the described examples has GU-AG intron boundaries at putative splice sites, and although a non-conventional splicing processing could explain the produced molecule, another type of RNA processing could not be ruled out. Processing of independently transcribed RNA from closely linked genes, that obey the minimal features to be considered as trans-splicing, presents lower frequencies when compared to the cis-splicing counterpart, but the protein products translated from the trans-spliced molecules have not been detected. The lower frequencies and the scarce data of proteins produced by the trans-spliced mRNAs lead to the hypothesis that the phenomenon could be a noise in the cis-splicing of two different pre-mRNAs, transcribed in distinct sites, and eventually paired. This interpretation could explain the high frequencies of intragenic trans-splicing, since it is expected that splice sites in transcripts generated by the same factory have a higher probability of interaction. However, this hypothesis does not explain why intragenic trans-splicing is so uncommon or why, at least in one case, exon repetition is allele-specific. In this particular case, cis regulatory elements were proved to be important for the processing to occur. Although these elements and their mode of action were not determined, it is possible that they are restricted to a low number of alleles and, could be related to specific transcription and splicing characteristics. Exon duplication proved to be highly efficient but at least in one case it is not essential. In contrast, Drosophila trans-splicing proved to be efficient and essential, i.e. it is a form of expanding protein variability with a functional biological role. Frequencies of lola interallelic trans-splicing are very high when compared to the mammalian intergenic trans-splicing, but a little higher or similar when compared with intragenic trans-splicing. This observation suggests an effect of proximity and attests the above-mentioned idea that the closer two transcription and/or splicing factories are the more likely their respective pre-mRNAs are of being trans-spliced. Following this hypothesis, we could consider trans-splicing as a possible me-chanism to generate protein diversity in metazoa and that Drosophila consolidated the process by coupling it into a different mode of transcription where the homologous chromosomes are still paired. In spite of the examples described up to now, it remains difficult to assign a biological function for trans-splicing. A clearer scenario occurs for by SL-addition trans-splicing studies. One function of SL-addition trans-splicing is the individualization of messages from polycistronic mRNAs in a concerted action with poly-adenylation. This type of processing is essential in organisms that have genes organized in operons, like C. elegans, or have long polycistronic units, like trypa-nosomatids. This regulatory role is extreme in trypanosomatids where transcription rates of different genes are very similar and transcriptional units may be as large as half of a chromosome. Another function of SL-addition trans-splicing is related to the capping of the RNA molecule. Thus it could be considered as a trans-capping reaction connected to cytoplasm addressing, mRNA stability and interactions with the translation machinery. Recently, the discovery of alternative SL-addition trans-splicing in trypanosomatids raised the possibility of regulating gene expression by generating mature transcripts with different 5' and 3' UTRs. The impact of alternative trans-splicing was extended since alternative SL addition trans-splicing could occur inside ORFs resulting in different protein products. It is worth mentioning that SL-addition trans-splicing has been considered as a general mechanism required for the production of functional mature transcripts, while mammalian trans-splicing is proposed to be involved in generating protein diversity. It could be the case that different functions have evolved from a common chemical language of two transesterification reactions and then adjusted to specific modes of gene expression. Some organisms like Drosophila and trypanosomatids developed efficient strategies in utilizing trans-splicing as a source of gene expression. Eukaryotic viruses, which speak this same language, could have utilized it to interfere in cellular growth and death by refining some language structures, their cryptic sites. However, it is not possible to exclude completely the proposition that mammalian trans-splicing and alternative trans-splicing of trypanosomatids could represent a noise reaction with no associated biological function. Acquisition of more data is important for the discrimination of trans-splicing as simple side reactions from part of the gene expression repertoire that once was a noise but now is a bang for certain organisms. ACKNOWLEDGEMENTS To Dr Ariel Mariano Silber and Dr Carlos Eduardo Winter for the critical reading of the manuscript and relevant suggestions. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05122f1.jpg] [oc05122f2.jpg] [oc05122f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}