 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 5, August 2005, pp. 555-561 Participation of cytokines in the necrotic-inflammatory lesions in the heart and skeletal muscles of Calomys callosus infected with Trypanosoma cruzi Isis F Magalhães-Santos, Sonia G Andrade/+ Laboratório
de Chagas Experimental, Auto-imunidade e Imunologia Celular, Centro de Pesquisas
Gonçalo Moniz-Fiocruz,
Rua Valdemar Falcão 121, 40295-001 Salvador, BA, Brasil Financial support: Fapesb, process 19.571.049.3383 Received 1 February
2005 Code Number: oc05129 Calomys callosus, a sylvatic reservoir of Trypanosoma cruzi, when infected with the Colombian strain (Biodeme Type III, T. cruzi I ) develops necrotic-inflammatory lesions and intense early fibrogenesis in the heart and skeletal muscles, that spontaneously regress. Participation of pro-inflammatory and pro-fibrogenic cytokines, such as tumor necrosis factor-alpha (TNF-α), gamma interferon (IFN-γ) , and tumor growth factor-beta (TGF-β), in the pathogenesis of the lesions is herein studied. Eighty C. callosus weighing 20 to 30 g were used. Seventy of them were inoculated with the Colombian strain (105 blood forms) and 10 were maintained as intact non-infected controls. After infection, C. callosus were sacrificed at different time-points from 15 to 70 days. The heart and skeletal muscle were processed for histopathology and cryopreserved for immunohistochemistry. Early necrotic lesions of parasitized skeletal muscle and myocardium with intense inflammatory lesions were present. Search for the in situ presence of TNF-a and IFN-γ, was performed using rat-IgG anti-mouse antibodies against these cytokines. For the in situ search of TGF-β, rabbit IgG anti-mouse antibodies were used. Immunolabeling of the cytokines in tissues of infected C. callosus was successful. The cytokines TNF-α, IFN-γ , and TGF-β were detected in the cytoplasm of macrophages and in the necrotic material from 15 to 45 days post-infection, decreasing their intensity until complete disappearance by the 65th day, which correlated with subsiding histopathological lesions. These findings suggest the participation of these cytokines in the control of parasite multiplication, in the development of an early fibrogenesis and in the regression of fibrotic-inflammatory lesions observed in C. callosus. Key words: Trypanosoma
cruzi - Calomys callosus - cytokines - tumor necrosis factor-alpha
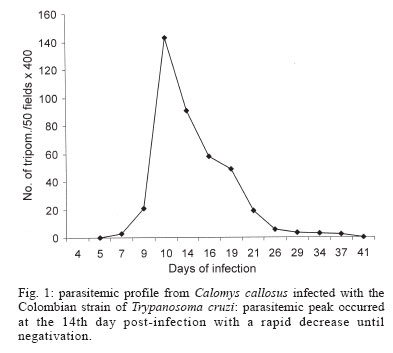
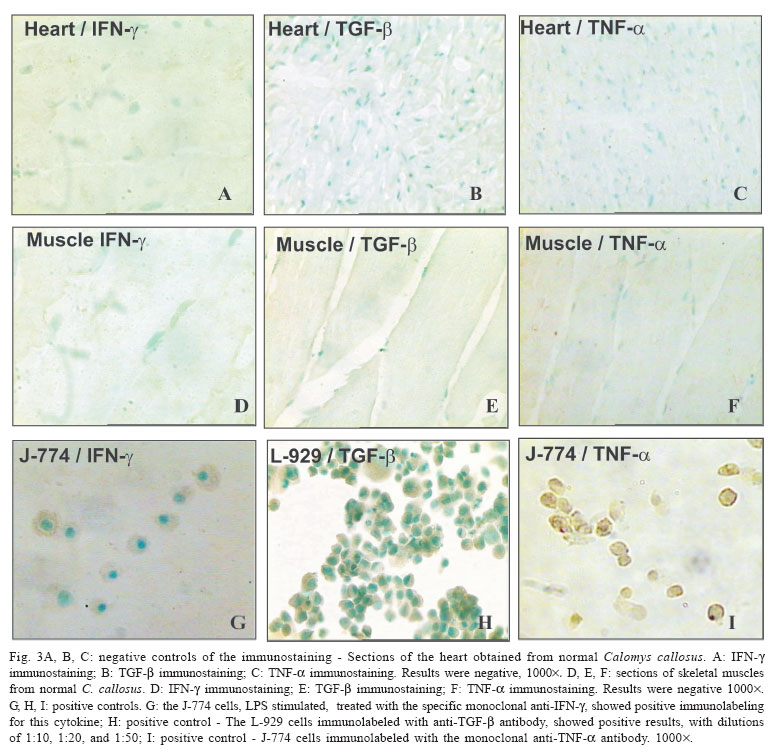
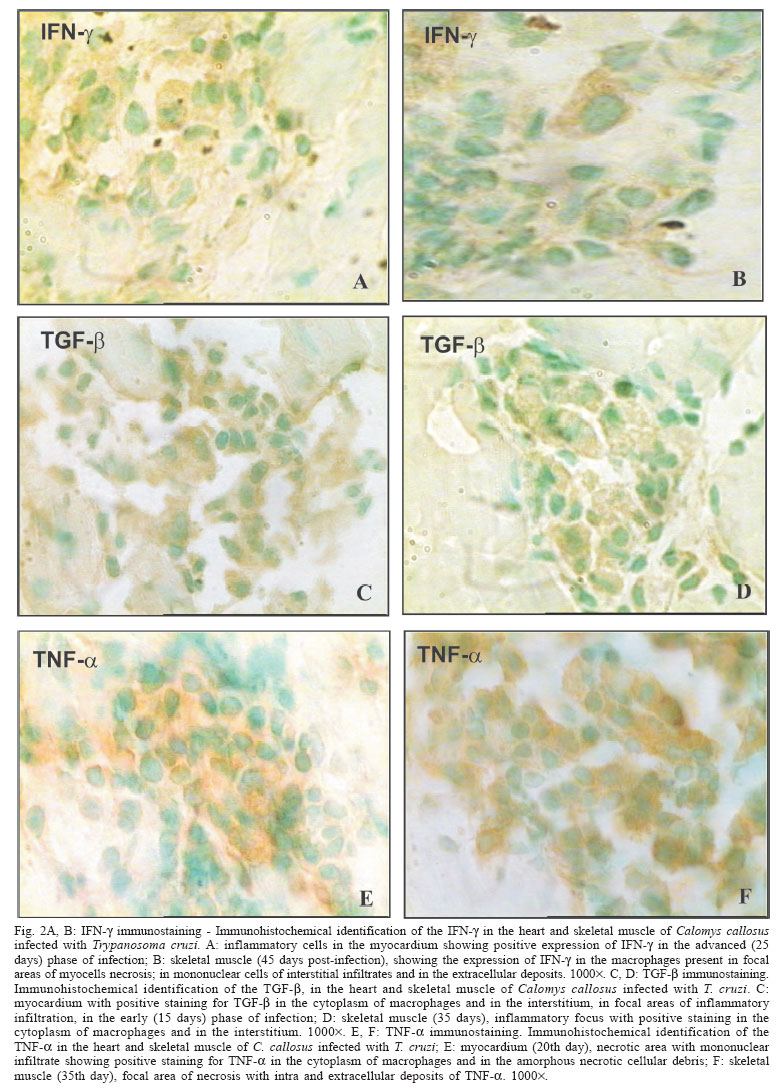
- gamma interferon - The Calomys callosus (Rodentia cricetidae) is a sylvatic vertebrate, natural reservoir of Trypanosoma cruzi. Adapted to laboratory studies (Mello 1982) this animal represents a very peculiar experimental model, with the capacity of to control the infection with virulent strains of T. cruzi as the Y strain (Mello 1979, Borges et al. 1982, 1992). Interestingly, tissue lesions are precocious with intense inflammatory infiltrates and early fibrogenesis, with a spontaneous regression of inflammation and fibrosis (Andrade et al. 1994, Lenzi et al. 1995, Magalhães-Santos et al. 2002, 2004). The basic mechanisms involved on the pathogenesis of the lesions and their regression are still unknown. It is probable that pro-inflammatory, fibrogenic, and regulatory cytokines play an important role. Thus, it appears important to investigate their participation in cardiac and skeletal muscles lesions in the host C. callosus, for the understanding of the immunological responses. In the C. callosus infection it has been demonstrated that parasite strains, especially those belonging to Biodeme Type III (T. cruzi I), as the F strain (Andrade et al. 1994) and the Colombian strain (Magalhães-Santos et al. 2002) are crucial for the determination of tissue lesions, since they are the most pathogenic. However differently from the mouse, the tissue lesions in C. callosus are progressive, going from the acute phase of infection and spontaneously subsiding by the early chronic phase (Magalhães-Santos et al. 2002). Studies with the murine model have shown that cardiomyocytes when infected with T. cruzi can produce marked amounts of cytokines such as gamma interferon (IFN-γ), interleukin 1-beta (IL-1β), tumor necrosis factor-alpha (TNF-α), interleukin 10 (IL-10), and tumor growth factor-beta (TGF-β) that can induce or regulate the production of different chemokines that modulate cellular immune response (Teixeira et al. 2002). In previous investigations Borges et al. (1995) have demonstrated the increase of IFN-γ levels in the serum of C. callosus acutely infected with a sylvatic strain of T. cruzi (M226). In the infection with others strains of T. cruzi, the levels of serum IFN-γ were high in the acute phase, with positive correlation with H2O2 liberation by peritoneal macrophages and it was associated with the decrease of parasitemia (Borges et al. 1995). The referred authors concluded that during the acute phase of the infection, the C. callosus develops a T-cell dependent immune response characterized by the production of IFN-γ with regulation of parasitemia to low levels, and controlling tissue parasitism without resulting in host-tissue aggression. Several immunological aspects, such as cytokines production in situ remain to be investigated to explain the spontaneous capacity of C. callosus to control the infection after significant tissue lesions caused by the presence of T. cruzi. In the present study, the in situ investigation of TNF-α, IFN-γ, and TGF-β revealed their presence in the necrotic-inflammatory lesions of the myocardium and skeletal muscles. These findings suggest the participation of these cytokines in the intracellular destruction of parasites, with evident control of parasitemia. The presence of necrotic inflammatory lesions, correlate with the high expression of TNF-α and IFN-γ. The presence of TGF-β coincides with regression of inflammation and degradation of fibrosis. This seems to indicate that these several factors are combined in the evolution of infection in the C. callosus. MATERIALS AND METHODS Experimental animals - Eighty C. callosus raised in the animal facilities of Centro de Pesquisas Gonçalo Moniz- Fiocruz, were used in this study. The experimental animals were approximately 60 day old males, weighing 20-30 g.Their maintenance complied with the guidelines for the human use of laboratory animals. Inoculation with T. cruzi - Seventy C. callosus were intraperitoneally inoculated with the Colombian strain (Biodeme Type III, T. cruzi I). Inoculum: 105 blood forms obtained from infected mice, after washing with PBS, pH 7.2, by centrifugation at 1200 g. Ten non-infected C. callosus were used as intact controls. Parasitemia of the infected animals was evaluated by counting trypo-mastigotes in 50 microscopic fields (400´) in peripheral blood, examined under slide and coverslip, three days/week. Histopathology - Five infected C. callosus per point were sacrificed with 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, and 70 days after infection. The heart and thigh-muscle fragments were collected and divided into two parts, one fixed in buffered formalin, pH 7.6 and the other cryopre-served for immunohistochemistry. For histopathology, tissues were embedded in paraffin and 5 µm-thick sections of the heart and skeletal muscle were stained with hematoxilin eosin and Picro-Sirius red method for collagen (Junqueira et al. 1979). Normal controls were sacrificed at the end of the experiments and submitted to the same procedures Immunohistochemistry - Fragments of heart and skeletal muscle were immediately embedded in tissue tek (OCT compound Miles Inc. Diagnostic Division, Elkhart, US) frozen in liquid nitrogen and cryopreserved in a freezer at 70oC until use. Sections of 5 µm obtained in a cryostat at -10oC placed on slides previously treated with 10% Poly L-Lisin (Sigma, St. Louis, MO, US) fixed in PA acetone were treated with PBS pH 7.4, containing 0.1% saponin (Sigma) and 1% bovin serum albumin (BSA) (Sigma). For blocking non-specific bindings, sections were treated with 10% BSA for 20 min at room temperature. As primary antibodies, the following antibodies were used in three experiments: (1) rat IgG antimouse TNF-α monoclonal antibody (MP6-XT3) (Pharmigen, San Diego, CA, US); (2) rat IgG antimouse IFN-γ monoclonal antibody (Sigma); (3) rabbit anti-mouse TGF-β purified polyclonal antibody (TGF-β 1, b 2, b 3, and b 5) (R&D Systems Inc. Minneapolis MN, US - Cod: DK-20-PB). Sections were incubated overnight at 4oC in an humidified chamber, with each antibody diluted in PBS pH 7.4/saponin 0.1%/BSA 1% in the dilution of 1:20 for anti-TNF-α and anti-IFN-γ; 1:50 for anti-TGF-B. After washing with PBS and saponin, sections treated with TNF-α or IFN-γ were incubated in normal rat serum in the dilution of 1:20 in PBS/BSA (5%) for 20 min, for additional blocking of nonspecific binding. For TGF-β the additional blocking was done with normal rabbit serum, diluted 1:20 in PBS/BSA (5%), incubated during 20 min. Inhibition of the endogenous peroxidase was achieved by a 30 min incubation in 0.3% H2O2 at room temperature. The slides were then incubated for 30 min at 37o C in a humidified chamber with the secondary antibody conjugated to peroxidase. For TNF-α and IFN-γ detection, a sheep anti-rat IgG antibody conjugated to peroxidase (Boehringer-Mannheim Biochemica, Germany) in the dilution of 1/300 in PBS/BSA/saponin, was used. For TGF-β a goat anti-rabbit peroxidase (Sigma) conjugate in the dilution of 1/800 in PBS/BSA/saponin was used. The colour was developed with 0.6 mg/ml 3,3' diaminobenzidine tetrahydrochloride (DAB) (Sigma) and 0.1% H2O2 plus 1% dimethylsulphoxide (DMSO) (Sigma). Sections were counterstained with 1% methyl-green for 15 min, dehydrated and mounted with permount. Positive controls - As control of the reaction for TNF-α and IFN-γ identification cells of the macrophage lineage J-774 (Ralph & Nakoinz 1975), described as "macrophage like" were used. The cells were maintained in RPMI cell culture medium with 10% bovine foetal serum, and collected during the exponential phase growth, at a concentration of 106cells/ml. The cells were stimulated for 24 h with 10 ng/ml with lipopolysacharide (LPS) from the cellular membrane of Escherichia colli (Sigma) in a CO2 chamber, at 37oC. Stimulated cells were centrifuged at 1500 rpm for 10 min. The pellet was suspended in PBS at a concentration of 106 cells/ml and re-centrifuged at 500 rpm for 5 min in a cytocentrifuge (Cytospin-3-Shandon, Life Sciences International, England) for adhesion to slides previously treated with Poly-L-lysine. As control of the reaction for TGF-β, L-929 fibroblasts from mouse (Willingham & Pastan 1974), were used. The cells were maintained in RPMI cell culture medium with 10% bovine foetal serum, and collected during the exponential phase growth, at a concentration of 106 cells/ml. These cells without stimulation, were collected, fixed in PBS + paraformaldehyde and processed in the same way as for the J-774 cells. Negative controls - (1) for each section of heart and skeletal muscle from infected animals a correspondent section, obtained from normal intact animal was processed through all steps of immunolabelling (to discard the possibility of cross reaction with C. callosus tissue proteins); (2) sections of the tissues either from infected or normal controls were performed through all steps, excluding the primary antibodies anti each cytokine and maintaining the secondary antibody (rat IgG peroxidase conjugate), to discard the possibility of unspecific reactions. RESULTS Parasitemia - Fig. 1 shows the profile of parasitemia, with peak at the 14th day post-infection. Histopathology - As previously described (Magalhães-Santos et al. 2004), histopathological lesions are prominent in the heart and skeletal muscles, in C. callosus infected with the Colombian strain of T. cruzi. Summarizing, intense inflammatory infiltration in the heart and skeletal muscles and necrotic lesions of the parasitized myocells were present from the 15th to 20th days of infection, predominantly in the myocardium. Cardiac lesions decrease in intensity from 25 to 35 days, being represented by focal necrosis of myocells and focal mononuclear infiltrations. Mild fibroblasts proliferation and interstitial matrix deposits were present. Regression of inflammatory and fibrotic lesions occurred from the 40th day of infection. Lesions of the skeletal muscle, increased in intensity from 25 to 30 days of infection, with necrosis of parasitized muscle cells, presence of disintegrated polymorphonuclear neutrophils and parasite debris, macrophages, and fibroblasts. Regressive lesions with focal mononuclear infiltration, were present from 35 to 45 days; fibroblasts proliferation and collagen deposit appeared in the interstitium as slender bundles. Parasitism was scarce or absent. Normal controls - Cardiac and skeletal muscle sections showed characteristic histopathological structure. Immunohistochemistry - Cross-reaction of the anti-mouse antibodies with the tissues of C. callosus was detected. The monoclonal antibodies anti TNF-α, IFN-γ, and TGF-β revealed specific immunolabelling of macrophages and necrotic tissues. Normal controls - Sections of the heart and skeletal muscle of intact C. callosus treated with each monoclonal antibody through all steps of the procedures to im-munolabeling of the cytokines: TNF-α, IFN-γ , and TGF-β, showed negative results. IFN-γ immunolabelling - Positive control: LPS stimulated J-774 macrophages were used to reveal the presence of IFN-γ in the cytoplasm of macrophages, taking into account that macrophages are able to release this cytokine during mycobacterial infection (Fenton et al. 1997, Swian 2001). As such, stimulated J-774 cells presented positive immunostaining for IFN-γ (Fig. 3G). Heart and skeletal muscle of infected C. callosus: from the 15th to 25th day post infection, sections of the heart showed positive immunolabelling for IFN-γ in the cytoplasm of macrophages and in extracellular material, corresponding to the inflammatory infiltrates around necrotic myocells (Fig. 2A) and in the extensive areas of necrosis of the skeletal muscle. With 35 and 45 days, the myocardium and the skeletal muscle showed focal and interstitial infiltrates with positive deposits in the cytoplasm of the mononuclear cells and in the interstitium (Fig.2B). By the 65th days post infection, staining was negative for IFN-γ in the myocardium. However, the skeletal muscle showed focal inflammatory infiltrates with positive staining for IFN-γ. TGF-β immunolabeling - Positive control: the L-929 cells immunolabeled with anti-TGF-β antibody, showed positive results (Fig. 3H), with dilutions of 1:10, 1:20, and 1:50. Heart and skeletal muscle showed in the early (15th and 20th days) and advanced phases (35th to 45th days) of infection, TGF-β was detected in the myocardium in the cytoplasm of macrophages and fibroblasts; positive extracellular deposits, were also present, in focal inflammatory foci around necrotic myocells (Fig. 2C). In the skeletal muscle, deposits of TGF-β were seen as positive mild staining in the cytoplasm of macrophages and fibroblasts and in extracellular material, from the 15th to 45th day of infection (Fig. 2D). However, TGF-β was not detected in the amorphous material of the extensive necrotic-inflammatory lesions in the skeletal muscle, present in the 25th day of infection. In the 45th days of infection, rare cells with scarce positive staining for TGF-β were present in focal inflammatory lesions. By the 65th days, residual infiltrates were seen in the myocardium and skeletal muscle, TGF-β being absent. TNF-α immunolabeling - As a positive control for TNF-α antibody, the J-774 cells were immunolabeled with the monoclonal anti-TNF-α antibody with positive staining (Fig. 3I). Sequential study from the 15 to 25 days of infection, showed the expression of TNF-α in small inflammatory foci in the myocardium, corresponding to areas of focal necrosis of myocells (Fig. 2E), in the cytoplasm of macrophages and in the amorphous necrotic cellular debris. Intense positive immunolabeling for TNF-α in the cytoplasm of macrophages and in the extracellular necrotic material was present mainly in the skeletal muscle. From the 35th to the 45th day of infection, small foci of inflammation with positive intra and extracellular deposits of TNF-α, were seen in the myocardium and skeletal muscle (Fig. 2 F). At 65th days of infection, residual infiltrates were seen in the myocardium and skeletal muscle, but these deposits does not present positivity for TNF-α. DISCUSSION T. cruzi infection determines parasitism of the myocardium, with intense myocarditis due to an immunological response at cellular and humoral levels, with production of a panel of inflammatory and regulatory cytokines (Gazzinelli & Brener 1991, Morgan et al. 1996) as seen in human patients from endemic areas, during acute infection. Experimental investigations of immunologic responses during acute infection have preferentially been developed in the mice model (Silva et al. 1991, Abrahamsohn 1998) which contributes to clarify the pathogenesis of lesions. In the murine model, infection with T. cruzi is followed by activation of macrophages as an initial innate response, mediated by IFN-γ produced by natural killer cells (Cardillo et al. 1996). Macrophages are then primed for further production of cytokines as TNF-α, TGF-β, and others, and to produce microbicidal agents such as N and O metabolites: nitric oxide (NO) and H2O2 (Gazzinelli et al. 1992, Silva et al. 1995, Dos Reis 1997). In the model of C. callosus, the investigation on the participation of the different cytokines in the progressive lesions has been restricted by the absence of available specie-specific monoclonal antibodies. In previous studies (Borges et al. 1995), evaluation of IFN-γ levels in C. callosus infected with T. cruzi, was performed by titration of antiviral activity on L-929 cells, by using a standard encephalomyocarditis virus cytopathic-effect-inihibiton. To characterize the antiviral activity, serial dilutions of serum samples were incubated with anti-murine IFN-γ monoclonal antibody. The antiviral activity has been tested both in C. callosus and mouse sera and was abolished 100% by the monoclonal antibody. The neutralization effect implies in a cross-reactivity between IFN-γof mice and that of C. callosus. Cross-reactive antibodies to mouse cytokines have been used in the present study, for the demonstration of the cytokines involved in the different phases of infection. In situ demonstration, of the cytokines involved in the response to infection with T. cruzi, has been done by Zhang and Tarleton (1996), in the spleen of infected mice. Pro-inflammatory cytokines as TNF-α and IFN-γ, as well as the regulatory cytokine TGF-b were identified during the course of infection in this study, and persisted until an advanced phase (70 days) (Zhang & Tarleton 1996). It has been demonstrated histochemically (Lima et al. 2001), in highly susceptible inbred mice (C3H/ He), infected with a macrophagotropic strain (Biodeme Type I), the participation of TNF-α in the massive parasite destruction and spleen necrosis. In mice model it has been demonstrated that macrophages of susceptible inbred mice produce higher levels of TNF-αthan the most resistant strains (Russo et al. 1989). Differing from the murine model, the evolution of the lesions in C. callosus infected with the Colombian strain was very rapid, corresponding to an early elevation of parasitemic levels, with peak at the 14th day, followed by a drop in crisis. This means that the C. callosus is highly susceptible to T. cruzi infection but controls the parasite multiplication. These observations were associated with in situ expression of TNF-α and IFN-γ. Probably the TNF-α and IFN-γ are mediating parasite disintegration by macrophage activation, with release of O metabolites such as H2O2, as demonstrated by Borges et al. (1995). In the present study it has been demonstrated that IFN-γpersists in C. callosus tissues for a prolonged time (65 days) as compared with the other cytokines that were absent in the heart and skeletal muscle at this phase. Serum levels of IFN-γ were evaluated in C. callosus and Swiss mice, infected with different strains of T. cruzi (Borges et al. 1995), in an attempt to correlate this cytokine with the control of infection in vivo. The highest IFN-γ serum levels in C. callosus, were obtained with strain F (Biodeme Type III, T. cruzi I), with peak at 7-day and with titers maintaining a plateau until day 28 and dropping after that to baseline levels. Differing from C. callosus, in the infection of mice with the F strain (Borges et al. 1995), IFN-γ peaks occur lately, between days 21 and 28 after infection, with a retarded drop. According to Silva et al. (1991) the regulation of IFN-g mediated killing of the intracellular T. cruzi amastigotes in mice, is mediated by TGF-β produced during acute infection. In vivo studies (Silva et al. 1991) have shown that the administration of TGF-β on T. cruzi infected resistant mice led to significantly increased parasitemias and mortalities. It is also a potent regulator of inflammation, being pro-inflammatory at low concentrations and anti-inflammatory at high concentrations (Omer et al. 2000). According to these authors T. cruzi is unable to invade cells lacking TGF-β receptors I or II. In the C. callosus model, the presence of TGF-b in the lesions of myocardium and skeletal muscle was detected since the early phase. This is consistent with the participation of TGF-b in the modulation of the other cytokines and in the early fibrogenesis, which follows the repair of lesions. The exsudative lesions characteristic of the intracellular parasites disintegration are replaced by macrophages collections and epithelioid cells, forming granulomatous lesions and proliferation of fibroblasts, characteristics of the delayed cellular immune response. Stimulation of fibrogenesis became clear, coincident with the presence of TGF-β, a pro-fibrogenic and regulatory cytokine in the cytoplasm of the macrophages and fibroblasts (Varga et al. 1987, Branton & Kopp 1999). The decrease and disappearance of the tissue cytokines, from 35 to 45 days of infection coincided with the regression of the inflammatory and fibrotic lesions, although with persistence of IFN-γ until later on (65th day). The results of the present study suggests that the in situ expression of cytokines such as IFN-γ, TNF-α, and TGF-β, is associated with the controlling of the parasite multiplication and precocious modulation of tissue lesions in C. callosus infected with T. cruzi. These findings demonstrated that this sylvatic animal is highly susceptible to the infection with the Colombian strain (Biodeme Type III, T. cruzi I). However it has the capacity of controlling the parasitic proliferation, without dying, a very convenient arrangement for a natural reservoir of parasites. ACKNOWLEDGMENTS To Dr José Mengel, who provided the anti-TGF-β purified polyclonal antibody. To Dr Fabiola Cardillo for reading the text and valuable suggestions. To Biologist Ariane R Pimentel to her assistance in the development of the immunohistochemical techniques. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05129f2.jpg] [oc05129f1.jpg] [oc05129f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}