 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 100, No. 7, November ,2005, pp. 693-698 Application of molecular techniques in the study of Staphylococcus aureus clonal evolution - A Review Adriana Marcos Vivoni, Beatriz Meurer Moreira+ Instituto de Microbiologia
Professor Paulo de Góes, CCS, Bloco I, Universidade Federal do Rio
de Janeiro, 21941-590 Financial suport:
CNPq, Capes, Faperj, the Fogarty International Program in Research and Training
in Emerging Infectious Diseases (TW006563) NIH Received 8 July
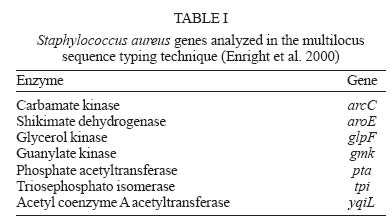
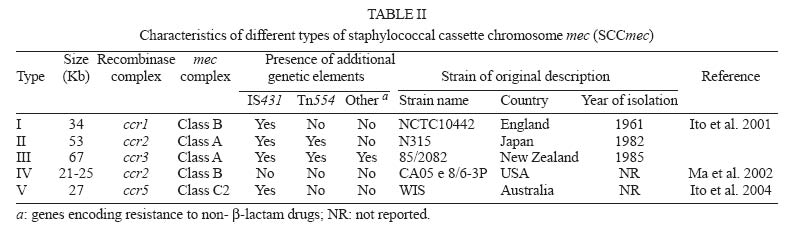
2005 Code Number: oc05146 Staphylococcus aureus is an important agent of healthcare-associated and community-acquired infections. A major characteristic of this microorganism is the ability to develop resistance to antimicrobial agents. Several molecular techniques have been applied for the characterization of S. aureus in epidemiological studies. In the present review, we discuss the application of molecular techniques for typing S. aureus strains and describe the nomenclature and evolution of epidemic clones of this important pathogen. Key words: Staphylococcus aureus - pulsed field gel electrophoresis - multilocus sequence typing - staphylococcal cassette chromosome mec typing - epidemiology During the last decade, Staphylococcus aureus was recognized as the most frequent agent of bloodstream and skin infections in Latin American hospitals (Diekema et al. 2001). Acquisition of resistance to several antimicrobial agents has been a major problem for the treatment of infections by this pathogen in the hospital and community settings. Resistance to β-lactam agents that resist hydrolysis of penicillinase, such as methicillin and oxacillin, is named "methicillin resistance" and is considered of great importance since oxacillin is the drug of choice for the treatment of S. aureus infections (Paradisi et al. 2001). The first cases of methicillin-resistant S. aureus (MRSA) were reported in the early 1960s, a few years after the introduction of methicillin in therapeutics (Jevons 1961). MRSA isolates produce a new penicillin-binding protein (PBP), called PBP2a or PBP2', with low affinity for β-lactam drugs, encoded by the mecA gene (Chambers 1997). MRSA is currently the most commonly identified antibiotic-resistant pathogen in US hospitals (NNIS 2004). Molecular typing techniques are widly applied in studies of S. aureus epidemiology, especially MRSA, with the aim of distinguishing isolates that are epidemiologically related from those unrelated. S. aureus outbreaks in hospitals are frequently considered as short duration events of localized epidemiology and, in these situations, most of the molecular typing methods are able to distinguish the isolates that belong to the outbreak (Shopsin & Kreiswirth 2001). However, the study of isolates that do not share a temporal or geographic origin, to define ancestors and lineages, is a major challenge. To date, several techniques allowed for the investigation and characterization of MRSA epidemic clones, but there are few data available about methicillin-susceptible S. aureus (MSSA). However, MSSA still causes about 50% of the healthcare-associated infections and almost all the staphylococcal community-acquired infections. Molecular typing techniques employed in the study of S. aureus epidemiology One of the first molecular techniques applied to study S. aureus epidemiology was the determination of the electrophoretic migration profile of plasmids in agarose gels. After the polymerase chain reaction (PCR) technique became available, several methods were developed and have been applied for S. aureus typing. PCR technology allows the evaluation of the structural organization of resistance determinants, such as the mecA gene, in isolates of different origins (Lim et al. 2002). In addition, some sequences can be used as targets for PCR-typing, such as the Tn916-Shine-Dalgarno (Netto dos Santos et al. 2001). A different approach is the repetitive extragenic palindromic-PCR (Rep-PCR), using primers that target repetitive extragenic palindromic DNA regions. This method has been used for typing MRSA (Del Vecchio et al. 1995) as well as MSSA (Netto dos Santos et al. 2001) isolates. Moreover, primers can target random DNA sites, such as in random amplified polymorphic DNA (RAPD) and amplified fragment length polymorphism (AFLP) techniques (Saulnier et al. 1993, Grundmann et al. 2002). However, to date, no appropriate criteria have been developed to interpret data obtained with these techniques. The first studies on the global molecular epidemiology of S. aureus A major progress in the understanding of the global epidemiology of S. aureus occurred after the development of multilocus enzyme electrophoresis (MLEE), a technique to analyze the electrophoretic mobility of housekeeping enzymes (Boerlin 1997). Interesting data were reported in 1992 by Musser and Kapur by the analysis of 254 MRSA isolates obtained in nine different countries of four continents, between 1961 and 1992. The authors reported that the mecA gene was inserted in isolates of different S. aureus lineages, suggesting that the acquisition of mecA was a horizontal transmission event that occurred several times in MSSA giving origin to different MRSA lineages (Musser & Kapur 1992). However, diverging results were published by Kreiswirth et al. in 1993. In this study, 472 MRSA isolates recovered from countries of four continents were analyzed by DNA restriction with ClaI followed by hybridization with probes for mecA and Tn554. All isolates could be divided into a limited number of mecA hybridization patterns, suggesting that mecA was most likely acquired by S. aureus only once, and all MRSA clones were descendents of this prototype strain. This hypothesis was accepted until the middle 1990s. Evolution of typing techniques and the charac-terization of MRSA epidemic clones The study of S. aureus epidemiology achieved a major advance with the application of pulsed field gel electrophoresis (PFGE). In this technique, bacterial DNA is immobilized into agarose plugs where endonuclease digestion is carried out. Electrophoresis is then run under an electric field that changes in pre-determined intervals, allowing for the separation of DNA fragments ranging from 10 Kb to 800 Kb (Olive & Bean 1999), while the conventional electrophoresis allows the separation of fragments ranging from 500 pb to 20 Kb. In 1995, Tenover et al. proposed a protocol for the interpretation of results obtained by PFGE. The interpretation system was based on results obtained by the application of PFGE to groups of isolates involved in well-characterized S. aureus nosocomial outbreaks. Accordingly, at first, the isolates presenting the most common pattern are defined as the outbreak strain. Then, the number of fragment differences in the electrophoretic patterns of all other tested isolates are compared to the outbreak strain, and the analysis is carried out, as follows: if no differences are found, isolates are considered part of the outbreak; with one to three differences (one genetic event), isolates are considered probably related to the outbreak strain; with four to six differences, isolates are considered possibly related to the outbreak strain; and finally, with more than six differences, isolates are considered as unrelated to the outbreak strain. Tenover and collaborator's criteria proved to be very useful, making PFGE suitable for epidemiological studies involving isolates from a restricted area, recovered during a three-month period, or even more, and have been successfully applied in several studies (Hilton et al. 2002, Montesinos et al. 2002, Wei & Chiou 2002). Although interpretation criteria for PFGE had been proposed for the analysis of isolates recovered in a period of up to three months, the technique has been effectively applied in several studies of global epidemiology. The combination of PFGE and the study of ClaI::mecA and ClaI::Tn554 polymorphism patterns led to the characterization of five major MRSA clones or lineages described bellow. These clones were named Iberian, Brazilian, Hungarian, New York/Japan and Pediatric, reflecting the geographic region where they were first identified or indicating a unique epidemiological characteristic. The combination of the polymorphisms ClaI::mecA type I and ClaI::Tn554 type E (I::E) characterizes the Iberian Clone (IC). This MRSA clone was first described in Spain, 1989 (Dominguez et al. 1994) and later on, in Portugal (Ferrari et al. 2002), Italy and United Kingdom (Mato et al. 1998), Germany (Witte et al. 1994), Belgium, Switzerland, France (Deplano et al. 2000), Poland (Leski et al. 1998), Check Republic (Melter et al. 2003), and United States (Roberts et al. 1998a). The Hungarian Clone (HC) is characterized by the combination III::B. It was first described as extensively disseminated in Hungarian hospitals (Sanches et al. 1998), and later on, reported in hospitals in Thailand (Crisóstomo et al. 2001). The New York/Japan Clone is characterized by the combination I::A and was first identified as the predominant clone in New York (Roberts et al. 1998b), New Jersey, Pennsylvania, and Connecticut hospitals in US (Roberts et al. 2000). It was also described in a hospital in Tokyo (Aries et al. 2000). The Pediatric Clone is characterized by the combination II::NH and was first described in 1992, in a pediatric hospital in Portugal (Sa-Leao et al. 1999). It was also reported in Poland (Leski et al. 1998), US (Roberts et al. 1998b), Argentina (Corso et al. 1998), Colombia (Gomes et al. 2001), and Brazil (Nunes de Melo et al. 2004). The Brazilian Clone (BC) was first described by Teixeira et al. (1995) as the combination III::B. However, Oliveira et al. (2002) described the same clone as XI::B and the HC as III::B. Nevertheless, in 1994, Sader et al. (1994) had already reported the occurrence of an endemic MRSA clone in eight hospitals in São Paulo city. This was named SP Clone and was probably the first report of the BC. However, differences between the PFGE protocol used by Sader et al. and the other authors mentioned above makes the comparison of their results impossible. Later on, the BC was reported in Portugal (de Sousa et al. 1998), Argentina, Uruguay, Chile (Aires et al. 2001), and the Check Republic (Melter et al. 2003). According to Oliveira al. (2002), the dissemination of MRSA clones in some wide geographic areas probably reflects their increased ability to cause infections, persist and disseminate from one region to another, including different continents. Although PFGE is an appropriate technique to study outbreaks, it is not sufficient for long-term studies or for studies of global epidemiology. In addition to the limitations of the interpretation criteria related to short time periods and restricted geographic areas, PFGE presents reproducibility problems. Different results are obtained when the technique is performed in different laboratories, even when standardized conditions are used (van Belkum et al. 1998, Olive & Bean 1999). In 2000, Enright et al. described, for the first time, the application of multilocus sequence typing (MLST) for the characterization of MRSA and MSSA isolates. MLST is a genotypic version of MLEE where isolates are typed by sequencing variable regions of housekeeping genes (Table I). Each new sequence is a new allele, and the combination of seven alleles forms the allelic profile or sequence type (ST). The results of MLST have been used to build a database that allows comparisons among S. aureus isolates from all over the world. This database can be accessed in the website http://www.mlst.net. A limitation of MLST is, however, the need of DNA sequencing, which is restricted to few institutions. Right after MLST was described for S. aureus typing, a new approach was proposed to study MRSA: the staphylococcal cassette chromosome mec (SCCmec) typing. The genetic element SCCmec, described by Katayama et al. (2000), is responsible for the mobilization of the mecA gene and harbors ccrA, ccrB or ccrC genes, which encode chromosome recombinases, and the mec complex. The proteins encoded by the ccr genes drive the excision and integration of the cassette in a specific site of the S. aureus chromosome (attBscc) located close to the origin of replication. The allotypes 1, 2, 3, and 4 were described for ccrA and ccrB, whereas only allotype 5 was described for ccrC (Ma 2004). In addition to the mecA gene, the mec complex carries other genetic elements. Three classes of the mec complex, presenting distinct structures, were described for S. aureus: class A (mecI-mecR1-mecA-IS431), class B (IS1272-DmecR1-mecA-IS431 ), and class C2 (IS431-mecA-DmecR1- IS431) (Ito et al. 2001, 2004). Therefore, the SCCmec can be further classified into types, according to the combination of the mec complex class and the recombinase allotype (Table II), SCCmec type I presents mec complex class B and ccr1; type II, mec complex class A and ccr2; type III, mec complex class A and ccr3; type IV, mec complex B and ccr2; and type V, mec complex C2 and ccr5. Recently, two possible additional SCCmec types have been identified among Australian community-acquired MRSA (O'Brien at al. 2004). The association of MLST and SCCmec typing supported a new nomenclature proposal for the MRSA epidemic clones. The lineages were classified according to the ST, resistance phenotype (MSSA, MRSA, VISA or VRSA) and SCCmec type. For example, the IC would be known as ST247-MRSA-I. The nomenclature for the major MRSA epidemic clones is shown in Table III. It is hoped that this nomenclature will replace the existing arbitrary designations providing a systematic classification of S. aureus clones (Enright et al. 2002). Studying S. aureus clonal complexes: establishing evolutive relationships among isolates In 2002, Enright et al. analyzed an international collection of hospital and community-acquired S. aureus isolates by MLST and SCCmec typing. The analysis revealed new information about clonal groups of S. aureus. Several clonal complexes (CC), composed by S. aureus isolates with the same ST or related STs (presenting at least five loci in common), were identified. Five major CCs included MRSA isolates: CC8, CC5, CC30, CC45, CC22. Interestingly, the comparison of the nucleotide sequences of MRSA and MSSA isolates included in CC8 allowed the establishment of a common MSSA ancestor. Different MRSA isolates would have originated from this MSSA ancestor by distinct events of mec cassette acquisition. These observations, in addition to other studies involving MLST, met Musser and Kapur's (1992) ideas about the horizontal transmission of mecA. Studies applying PFGE for MSSA typing have shown a large diversity of electrophoretic profiles among these isolates (Enright et al. 2000, Netto dos Santos et al. 2001). MLST of MSSA shows overlapping results: isolates presenting related PFGE profiles usually share the same ST or related STs ( Enright et al. 2002, Vivoni et al. 2004). In 2001, Crisóstomo et al. analyzed a MSSA collection from the 1960s, obtained in Denmark previously to the first MRSA isolation. The study allowed the identification of probable MSSA ancestors of the Archaic, Pediatric and NY/Japan MRSA clones (Crisóstomo et al. 2001). However, studies involving more recent MSSA isolates (Mahmood et al. 2002, Enright et al. 2002, Vivoni et al. 2004) were not able to delineate associations between specific clones or CCs and the community or hospital origin of the isolates. They were able to show the predominance of some CCs though, suggesting that some lineages might present a better pathogenic or adaptive potential. Among isolates from the United Kingdom obtained between 1997 and 1998, the CC 30 is predominant, followed by CCs 25, 8 and 1 (Enright et al. 2000, 2002, Mahmood et al. 2002) In Brazil, isolates belonging to CC1, CC 30 and CC 5 are prevalent (Vivoni et al. 2004). Conclusions A better understanding of the epidemiology of S. aureus contributes to the control, prevention and treatment of healthcare-associated and community-acquired infections. Molecular techniques are important tools in outbreak investigations and in global epidemiology studies, not only for S. aureus, but also for many other pathogens. However, the application of the appropriated technique is critical for the success of the study and production of accurate results. Several studies cited in this review reinforce the idea that the horizontal transfer of the mecA gene to certain MSSA isolates originated MRSA lineages to become pandemic clones. However, studies involving isolates from several countries are necessary to establish the existence of disseminated S. aureus. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz |

{kind=link}
{kind=link}
{kind=link}