 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
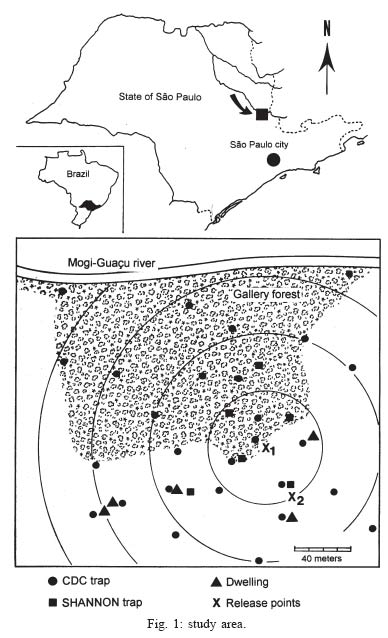
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 100, No. 7, November ,2005, pp. 719-724 Dispersal pattern of the sand fly Lutzomyia neivai (Diptera: Psychodidae) in a cutaneous leishmaniasis endemic rural area in Southeastern Brazil Cláudio Casanova/+, Antonio IP Costa, Delsio Natal* Superintendência de Controle de Endemias, Rua Afonso Pessini 86, 13845-206 Mogi Guaçu, SP, Brasil *Faculdade de Saúde Pública, USP, São Paulo, SP, Brasil Financial support:
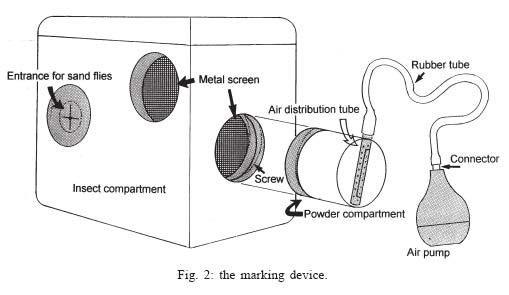
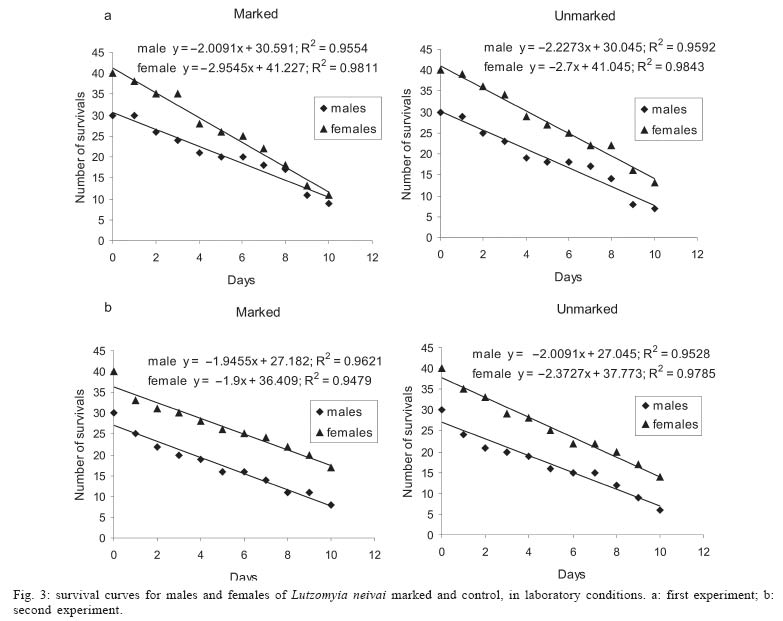
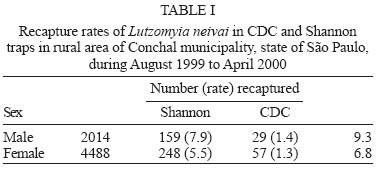
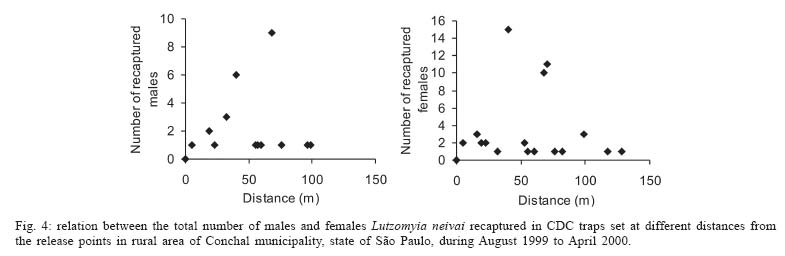
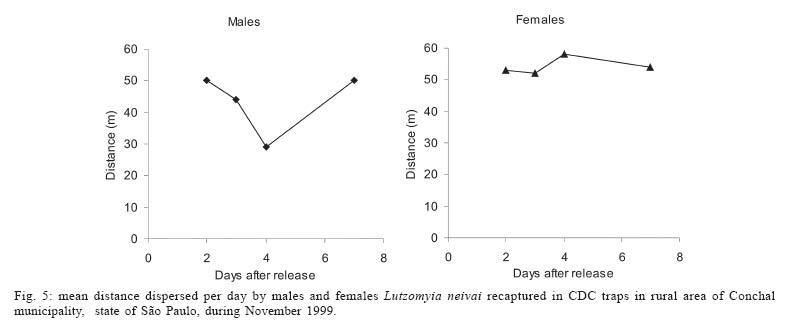
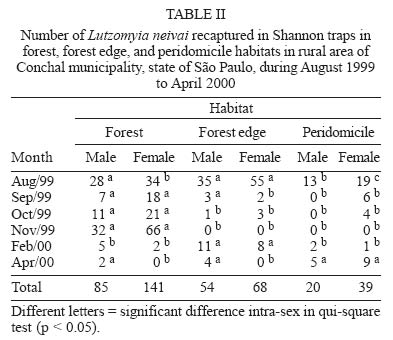
Sucen, CNPq Received 12 August 2005 Code Number: oc05151 The dispersal pattern of the sand fly Lutzomyia neivai was studied through mark-release-recapture experiments in an American cutaneous leishmaniasis endemic rural area in Southeastern Brazil. Over 6500 specimens were marked with fluorescent powder and released in forest edge and peridomicile habitats from August to November 1999, February and April 2000. Recapture attempts were made using Shannon and CDC traps up to eight successive nights after releases. A total of 493 (7.58%) specimens were recaptured. The number of recaptured males and females of L. neivai in CDC traps was not affected by the distance between the trap and the release points. Approximately 90% of males and females recaptured in CDC traps were caught up to 70 m from the release points. The maximum female flight range recorded was 128 m. The average flight range per day was less than 60 m for males and females. Of the flies released in forest edge, approximately 16% of the recaptured females were caught in Shannon traps in the peridomicile habitat. The results indicate that the movements of L. neivai are spatially focal and the possibility of dispersion from forest to peridomicile habitat may be an important way of contracting leishmaniasis in dwellings. Key words: mark-release-recapture - dispersal - sand fly - Lutzomyia neivai - Brazil Information on the dispersion pattern of sand flies vectors in nature can be used to evaluate the potential rate of Leishmania dissemination and to implement surveillance and control strategies against these vectors. Previous dispersal studies have shown that species from Old World arid and semi-arid regions may fly more than 1500 m (Killick-Kendrick et al. 1984, Doha et al. 1991), while sand flies from Neotropical forests seldom travel no more than 200 m from their release point (Chaniotis et al. 1974, Alexander 1987, Alexander & Young 1992, Morrison et al. 1993). In Brazil, the Lutzomyia longipalpis (Lutz & Neiva) dispersal patterns have been studied (Dye et al. 1991, Kelly & Dye 1997), but there is no study of the dispersal pattern of the vectors of American cutaneous leishmaniasis (ACL). In the Southeastern region of Brazil, the forest seems to be the environment of the enzootic cycle of Leishmania (Viannia) braziliensis (Gomes et al. 1989, Gomes 1994, Lainson et al. 1994), and it is mostly in the domiciliary habitat that man becomes infected (Rangel et al. 1990, Gomes 1994, Tolezano 1994, Gontijo et al. 2002). Thus the flight range and the dispersion of sand flies between these habitats are important factors in ACL transmission. The present study was carried out to describe the dispersal pattern of Lutzomyia neivai (Pinto, 1926), recently resurrected as species of the complex Lutzomyia in-termedia (Lutz & Neiva, 1912) by Marcondes (1996), in an ACL endemic rural area in Southeastern of Brazil, by the mark-release-recapture technique. The anthropophilic behavior of the species, its predominance in modified environments and its presence in houses were people have become infected indicates L. neivai as the suspected vector in a large area (Gomes 1994, Tolezano 1994, Casanova et al. 1995, Marcondes et al. 1998, Teodoro et al. 2001, Salomon et al. 2003, 2004, Andrade Filho et al. 2003). MATERIALS AND METHODS Study area - The study was carried out in an ACL endemic rural area in municipality of Conchal (22°18'S and 47°13'W), Mogi Guaçu River Valley, São Paulo (Fig. 1). This locality has a secondary remnant gallery forest, which covers the left margin of Mogi Guaçu river (Fig. 1), small farms with fruit-trees, vegetables, and domestics animals such as chickens and pigs. According to the Köeppen classification, the climate is subtropical warm with dry winter (Cwa). The average annual rainfall is 1381 mm and the average annual temperature is 21.6°C. Previous studies demonstrated that L. neivai is the predominant species in this area (Casanova et al. 1995, Casanova 2000). Mark-release-recapture experiments - Mark-release-recapture experiments were performed with wild-caught sand flies from August 1999 to April 2000. The sand flies were caught using the Castro aspirator (Alexander 2000) in modified Shannon traps (Vexenat et al. 1986) from 18 h to 21 h. The sand flies were counted and transferred to a Styrofoam cage (20×13×18 cm) with a thin layer plaster of Paris (Fig. 2) and then marked with fluorescent powder (red, white, yellow, and blue - Bioquip Inc.) according to the date and site. The powder was dispersed into the cage by puffing air in a separate powder compartment (5 cm diameter), similar to the one used by Pardo et al. (1996), which was connected by a hole (5 cm diameter) covered with a metal screen (0.40 mm2) to the cage. Another opening (2.5 cm diameter) covered with the same metal screen was made in the side of the cage to increase the air circulation during dusting (Fig. 2). The sand flies were released immediately after dusting from their capture sites on the edge of the forest. In two experiments, the flies were also released from a peridomicile site. Recapture attempts were performed for 5-8 successive nights after the release night using six Shannon traps and for 3-4 nights using 16-20 CDC traps. Two Shannon traps were set, from 18 h to 21 h, in forest, forest edge, and peridomicile habitats, between 25 m and 90 m from the release points (Fig. 1). The CDC traps were operated from 18 h to 7 h at 5 m to 200 m from the release points in the three habitats (Fig. 1). The material collected was taken to the laboratory and marked and unmarked sand flies were separated under a stereomicroscope and black-light-blue for fluorescence. All sand flies were prepared (phenol - potassium hidroxid - acetic acid - acid fucsin - etilic alcohol - and vegetal creosote) according to Forattini (1973) and identified according to Young and Duncan (1994). The number of marked individuals recaptured in each CDC traps was transformed to l n (y + 1) and regressed as a function of traps distance from the release points. The mean distance dispersed per day was estimated only for the November experiment, when the number of recaptured individuals in CDC traps was large enough. CDC trap collections were grouped into 40 m annuli from the point of release situated at the forest edge and adjusted by a correction factor, to accommodate unequal trap density, by the method of Brenner et al. (1984). The chi-square test was used to establish statistical differences in the proportions of males and females recaptured in Shannon traps between pairs in the forest, the forest edge, and the peridomicile habitats. Two laboratory experiments were carried out to determine the effect of the fluorescent powders on the sand flies' longevity. For each experiment, two lots of 30 males and 40 females of wild-caught L. neivai were transferred to two Styrofoam cages (20 ×12 ×15 cm) and one sample was dusted, as described above. Each sample was released in holding cages (20 × 20 cm) with 10% sucrose solution available and was maintained under laboratory conditions (26ºC and 82% RH). The number of dead males and females was recorded daily for 10 days. All sand flies were processed and identified. For statistical analysis, the untransformed number of males and females marked and unmarked of L. neivai was regressed as a function on time in days. Each regression coefficient was tested for significant departure from zero by ANOVA and comparison among regression coefficient were made by ANCOVA (Sokal & Rohlf 1995). RESULTS Laboratory experiments - In the control experiments (Fig. 3) the untransformed data provided a significant linear fit (p < 0.05) and the regression coefficient and adjusted mean did not differ significantly (p > 0.05) in the covariance analysis. In the second experiment, marked females lived longer than the control (ANCOVA < F1; 18 = 6.243; p = 0.021). Median life expectancies for marked and control were, respectively, 9 and 8 days for males and 8 and 9 for females in the first experiment and 7 and 6 days for males and 9 and 8 days for females in second experiment. The fluorescent powder was detected easily on individuals that died during the experimental period. Field experiments - A total of 6502 specimens were marked. The device used allowed the simultaneous marking of 1485 individuals. All the sand flies, 2014 males and 4488 females, marked and released were presumed to be L. neivai, since more than 99% of the sand flies collected in the attempted recaptures, and all the recaptured individuals belonged to this species. A total of 188 (9.3%) males and 305 (6.8%) females of L. neivai were recaptured (Table I). Approximately 85% of males and 81% of females were recaptured in Shannon traps. In these traps, the male recapture rate (7.9%) was significantly higher (χ2 = 11.9, df = 1; p < 0.005) than the female rate (5.5%) and in CDC traps these rates were similar (Table I). The number of recaptured individuals in CDC traps transformed in l n (y + 1) did not decrease significantly (for males: l n (y + 1) = -0.001x + 1.122; p > 0.75, and for females: l n (y + 1) = -0.003x + 1.414; p > 0.55) as a curvilinear function of trap distance from the release points (Fig. 4). The arithmetic mean distance dispersed by the males was 53 m and by the females 55 m and 86% of the males and 89% of the females were recaptured up to 70 m of their release points. The maximum dispersion distance was 99 m for males and 128 m for females that were recaptured 7 and 8 days after their release. The longest period between release and recapture was 11 days for females and 14 days for males released. The mean distance dispersed per day was calculated for 14 males and 45 females of L. neivai recaptured in the 16 CDC traps during the November experiments and was adjusted for the area sampled. Mean dispersal distances were practically stable between 40 and 60 m for males and females released at the forest edge (Fig. 5). Since the collection effort per habitat with Shannon traps was the same, it was possible to estimate the predominant direction of dispersion from the release point situated in forest edge based on the number of recaptured L. neivai. On three occasions for females (September, October, November) and two (October and November) for males there was a significant difference of dispersion in the Shannon traps situated in forest habitat, and on only one occasion for females (April) in the Shannon traps situated in peridomicile habitat (Table II). Pooling all the Shannon trap recaptures of females we found that 56.9% were from the forest, 27.4% were from the forest edge and 15.7% were from the peridomicile habitat (Table II). There was no evidence that L. neivai movement was influenced by the wind, because in all the experiments large numbers of recaptured sand flies were in Shannon traps situated in different sites that were independent of the predominant wind direction. DISCUSSION The comparable mortalities among marked and control groups of L. neivai in the laboratory experiments, coupled with the field recapture results, show that the marking device described in the present study can be safely used in other sand flies dispersal studies. Furthermore, this technique complies with one of the five basic assumptions of a mark-release-recapture method used to estimate the absolute size of population in nature (Southwood 1978) which is that the marked animal must not be affected by marking. The overall recapture rate of L. neivai (7.5%) was comparable with those reported by previous studies for other cutaneous leishmaniasis potential vectors in Neotropical region (Chaniotis et al. 1974, Alexander 1987, Alexander & Young 1992). The results of the arithmetic mean of dispersal distances (53 m for males and 55 m for females), the mean distance dispersed per day (bellow 60 m for males and females) and the maximum dispersal range (99 m for males and 128 m for females) indicates that the movements of L. neivai are spatially focal in our study area. These results agree with other sand flies dispersal studies in similar habitats of the Neotropical region. It is most probable, that the availability of diurnal resting sites, food sources, and breeding sites are in or near to the forest environment so that dispersion is not important (Chaniotis et al. 1974, Alexander & Young 1992). Although the marked individuals were not released for a second time during the field experiments, the recapture of two females and two males up to 11 and 14 days after release, respectively, represent the higher survival for Neotropical sand flies in nature. Estimates of daily survival during the gonotrophic cycle and the absolute abundance are fundamental in evaluating the vectorial capacity of L. neivai and will be addressed in a future article. The higher recapture of L. neivai females in Shannon traps situated in forest and forest edge (84.3%) environments demonstrate the tendency of this species to remain on the forest border or to disperse to forest environment. On the other hand, the fact that approximately 16% of recaptured females occurred in Shannon traps situated in the peridomestic habitat support the hypothesis that domiciliary transmission results from females infected in an enzootic cycles that is probably situated in the nearby forest (Gomes et al. 1989, Gomes 1994, Lainson 1994). Since the vector's dispersion is one of the major factors that determines the potential rate of pathogen dissemination, the focal dispersion of L. neivai suggests a low rate of L. (V.) braziliensis dissemination. Coupled to the dispersion to peridomicile habitat, the present data helps our understanding of the focal distribution of human cases and domiciliary transmission. This is the present epidemiological pattern of ACL in Southeastern region of Brazil in areas where L. neivai is the predominant species (Gomes 1992, 1994, Tolezano 1994). Besides this studies on the genetic structure of L. (V.) braziliensis populations have also indicated that they are focal (Ishikawa et al. 2002, Cupolillo et al. 2003) and that the reservoirs of this parasite also have focal habits (Brandão et al. 2003). The importance of the forest habitat and the dispersal potential of the vector provide important information that can be used to determine the feasibility of using "buffer zones" and their width, or the creation of vegetation-free belt between dwellings and the forest suggested by Alexander and Young (1992) as a potential alternative to chemical control used against the vectors. The recapture of one L. neivai female in a peridomicile at approximately 130 m from the release point situated in forest edge should be taken into consideration in the control activities against this suspected cutaneous leishmaniasis vector. ACKNOWLEDGMENTS To Dr Flávio AM dos Santos, Universidade Estadual de Campinas, for the statistical analysis. To Dr Jeffrey J Shaw for suggestions on the manuscript. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}