 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
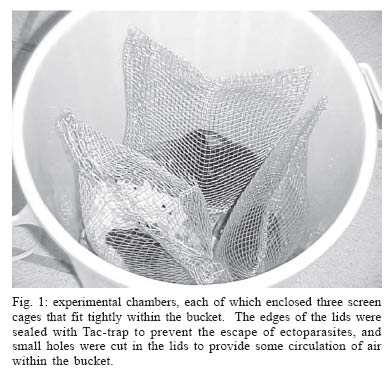
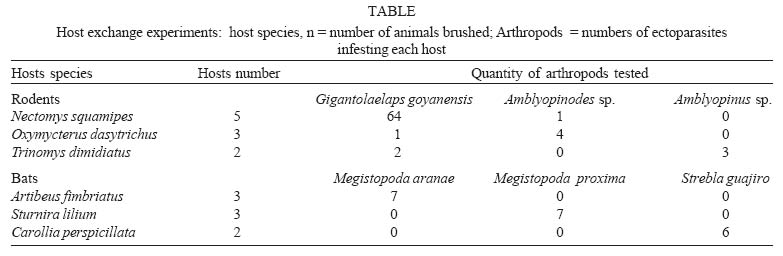
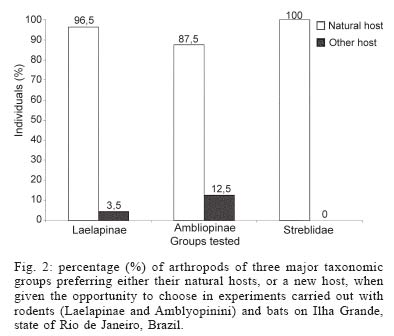
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 100, No. 7, November ,2005, pp. 761-764 A method for testing the host specificity of ectoparasites: give them the opportunity to choose Carlos EL Esbérard/+, Fernanda Martins-Hatano, Emerson B Bittencourt*, David EP Bossi**, Angélica Fontes, Marcela Lareschi***, Vanderlaine Menezes, Helena G Bergallo, Donald Gettinger**** Departamento de Ecologia, Ibrag, Universidade do Estado do Rio de Janeiro, Rua São Francisco Xavier 524, 20559-900 Rio de Janeiro, RJ, Brasil *Laboratório de Ixodides, Departamento de Entomologia, Instituto Oswaldo Cruz-Fiocruz, Rio de Janeiro, RJ, Brasil **Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis-Ibama, Brasília, DF, Brasil ***Centro de Estudos Parasitológicos y de Vetores, La Plata, Buenos Aires, Argentina ****Laboratory of Parasitology, Department of Biology, University of Central Arkansas, Conway, Arkansas, US Corresponding author. E-mail: cesberard@terra.com.br Received 12 July 2005 Code Number: oc05160 Host-choice experiments were carried out with rodent and bat ectoparasites on Ilha Grande, state of Rio de Janeiro, Brazil. We constructed experimental chambers that enclosed three different rodent or bat host species, and then introduced a selected set of ectoparasitic arthropods. When given the opportunity to choose among host species, the ectoparasites showed a strong tendency to select their primary hosts, and reject novel host species. These kinds of simple experiments can be valuable tools for assessing the ability of ectoparasites to locate and discern differences between host species, and make choices about which hosts to infest, and which hosts to avoid. Key words: ectoparasites - host-specificity - experiments - small mammals - neotropics In the opening address of the "First Symposium on Host Specificity among Parasites of Vertebrates" in Neuchatel, France, 1957 (Wenzel & Tipton 1966), Ernst Mayr posed a number of questions about host specificity that are still of great importance in the evolutionary biology of parasites and vertebrates. "Where does host specificity occur? How strict is it? What groups of parasites are most host specific? Why are some parasites highly specific, others of rather wide distribution?" Today, in studies of neotropical ectoparasites, the answers to these questions continue to be confounded by the same problems noted by Mayr and later lamented by Wenzel and Tipton (1966). An ectoparasite is considered host specific when it is associated with a single species of host (monoxeny) or a group of host species that are closely related phylogenetically (pleioxeny). Both vertebrates and their associated arthropods remain very poorly known in many parts of the neotropics, and most of our information is coming from large, extensive faunal surveys. Without a sufficient number of samples collected at the population level, the taxonomy of both vertebrates and ectoparasites is inadequate for many kinds of biological studies. When ectoparasites are collected from novel hosts, it can be the result of unreliable host or ectoparasite identifications, contamination of samples, or real biological events in ecological or evolutionary time. But survey data alone cannot be applied to the question of whether strict host specificity is due to an inability of the ectoparasite to survive on other host species, or simply an inability to locate and disperse to another host. An experimental approach is needed. Reed and Hafner (1997) conducted transfer experiments in the laboratory to determine if mallophagan lice can survive and reproduce on non-related gophers, and found that these permanent ectoparasites can, in fact, establish on other hosts. They concluded that host specificity was maintained by a lack of opportunity for lice to colonize new hosts. Tomkins and Clayton (1999) transferred host-specific mallophagan lice reciprocally among the nestlings of four species of cave swiftlets in the field in Borneo. Because the transferred lice did not survive well on new hosts, they concluded that even if lice could colonize new hosts, they could not acquire the resources for successful establishment. However, many ectoparasitic arthropods are more vagile than mallophagan lice, and may have better opportunities to come in contact with novel hosts. We wanted to develop an operational experimental strategy to evaluate the ability of ectoparasites to locate and discern differences between host species, and make choices about which hosts to infest, and which hosts to avoid. If host specificity occurs because of a limited capability to disperse among novel host species, when the barriers that impede them are removed, ectoparasites should colonize other hosts. However, if specificity occurs because of adaptive constraints, (i.e., a reduction of parasite fitness on atypical host species), then ectoparasites should prefer their typical hosts. The objective of this paper is to present a simple and original experimental method, easily replicated, for testing the host choice preferences of ectoparasitic arthropods. MATERIALS AND METHODS The experiments were carried out from November 8-14, 2002, in an area of Atlantic forest located on Ilha Grande, a large island off the mainland of the coast in the state of Rio de Janeiro (23° 05' to 23° 15'S, 44° 06' to 44° 23'W), at a research facility, Centro de Estudos Ambientais e Desenvolvimento Sustentável (Ceads) of the Univer-sidade do Estado do Rio de Janeiro A transect of 100-130 livetraps, and 5 or 6 mist nets set along an Atlantic Forest stream provided, respectively, a daily supply of rodents and bats. We constructed closed experimental chambers, where ectoparasitic arthropods were introduced and offered a choice of three different vertebrate hosts, which were placed in close proximity within the chambers, and connected by a common nest substrate (rodents) or roosting surface (bats). The chambers were plastic garbage buckets of 30 l capacity, each of which enclosed three screen cages (3/4" mesh) that fit tightly within the bucket (Fig. 1). The edges of the lids were sealed with Tac-trapÓ to prevent the escape of ectoparasites, and small holes were cut in the lids to provide some circulation of air within the bucket. Rodent experiments - Three small mammals common to the study area, two murid rodents, Nectomys squamipes (Brants 1827), Oxymycterus dasytrichus (Fischer 1814), and the echimyid rodent, Trinomys dimidiatus (Gunther 1877), were used in the experiments. During the early morning, rodents captured in the field were brought into the laboratory, placed in a small plastic box and anesthesized with ether. Each animal was brushed thoroughly with a toothbrush to remove the ectoparasites (see Martins-Hatano et al. 2002 for further information on ectoparasite collection). An artificial nest substrate made of strips of paper towels was placed in the bottom of the bucket, and in the bottom of the wire holding cages, providing easy access for crawling ectoparasites to move from nest to the rodent of choice. The removed ectoparasites were introduced into the bottom of the bucket, then the wire cages, each holding a different species of rodent (N. squamipes, O. dasytrichus, and T. dimidiatus), were placed into the bucket and the lid was secured. After 10-12 h, the rodents were removed from the holding cages, etherized, and brushed. The ectoparasites from each of the three experimental hosts were collected and placed in individual vials of 70% ethyl alcohol. Bat experiments - Three phyllostomid bats that are common in the study area, Artibeus fimbriatus Gray, 1838, Sturnira lilium (E. Geoffroy 1810), and Carollia perspicillata (Linnaeus 1758), were used in the experiments. Bat nets were opened at dusk and captured bats were transported to the laboratory within individual cloth bags, transferred into the small plastic box, and lightly etherized. The streblid flies (Megistopoda aranea (Coquillet 1899), Megistopoda proxima (Séguy 1926), and Strebla guajiro (Garcia & Casal 1965) were then collected manually and placed temporarily in small plastic vials. Each bat was thoroughly searched until all the flies were captured, then were given a sugar solution orally. The three experimental bat hosts (A. fimbriatus, S. lilium, and C. perspicillata) were placed in the buckets first, and then the captured flies were introduced through a hole in the top of the bucket. Because some streblids can fly (others are brachypterous and apterous), we covered the entrance hole with fine screening. After 10-12 h, the bats were removed from the holding cages, etherized, and brushed. The streblid batflies collected from the three experimental bats were placed in vials of 70% alcohol. Because we wanted to be sure that all the arthropod individuals were removed from the host both before and after the experiments, we chose to work with large arthropods. For rodents, we used laelapid mites of the genus Gigantolaelaps and amblyopinine beetles; with bats, we used streblid batflies. Previous studies of the rodent and ectoparasite fauna of Ilha Grande (Guitton et al. 1986, Martins-Hatano et al. 2002, Bittencourt & Rocha 2002) have provided evidence of strong specificity of these ectoparasites for their small mammal hosts. G. goyanensis is a host-specific ectoparasite of N. squamipes (also see Furman 1972, Gettinger 1987); Amblyopinus sp. is specific to T. dimidiatus; Amblyopinodes sp. to O. dasytrichus. For bats, we relied on the host records from large ectoparasite surveys in Venezuela (Wenzel 1976), and studies of other areas in the Atlantic Forest Region that have quantified the host associations of the arthropods utilized in our experiments (Bertola et al. 2005). Megistopoda aranae is a host-specific ectoparasite of A. fimbriatus; M. proxima is specific to S. lilium, and Strebla guajiro to C. perspicillata. RESULTS AND DISCUSSION Over a five day field trip, we carried out six experimental runs with the rodents and their ectoparasites, and two experimental runs with the bats and their ectoparasites (Table). All groups of ectoparasites consistently chose their natural hosts (i.e., the preferred host species as in the literature) (Fig. 2). In fact, they chose their natural hosts at about the same percentage as is registered in the literature (between 90 and 100% of the time). Hence, it was observed that most of the ectoparasite individuals, when given an equal chance to choose between the three hosts, returned to the original host species. These data support the larger body of evidence coming from extensive faunal surveys of neotropical ectoparasites and their mammalian hosts, and indicate that the described experiments can be used to evaluate the capacity of ectoparasites to distinguish among hosts in ecological time. New experiments could be performed, testing the preferences of ectoparasites for hosts of different sexes, age, body size, or reproductive activity, as well as preferences of different combinations of host taxa, perhaps even eliminating the primary, or natural host. These experiments can provide important supplementary and supportive information for both historical and ecological studies of vertebrates and their ectoparasitic arthropods. However, some suggestions can be made: (i) when utilizing ectoparasites of small size or with the ability to hold tightly to the host skin or pelage, we recommend that the host is brushed repeatedly to guarantee that all arthropods are removed before the host is placed into the holding cages (another possibility would be the use of insecticides to make sure the host is clean before experiments, but this would require a detoxification period); (ii) it may be possible to obtain counts of the number, sex, and/or life stages of the ectoparasites before placing them into the bucket, then using a Berlese funnel to determining the number of living arthropods left in the bottom of the bucket without choosing a host, and the number of arthropods that died during the experimental process (due to ether, manipulation, or grooming by the vertebrate hosts); (iii) following the host choice experiments, it may be possible to maintain the vertebrates infested with atypical ectoparasites in captivity to verify the capacity of these arthropods to survive and reproduce on novel hosts. Determining the host specificity of a parasite is a very complex problem, involving anatomical, physiological, evolutionary, and behavioral adaptations. Although these preliminary experiments are simplified, and we have employed only the most basic treatments (i.e., lumping laelapine mites and amblyopinine beetles in a single experiment), we believe that the approach is original and promising. For example, ectoparasites can also have spatial and temporal infestation preferences, choosing different regions of the host body at different times of the day (Bittencourt & Rocha 2002). By varying the experimental design, it may be possible to understand some of the complicating factors that determine host specificity. Experimental strategies have great potential for approaching ecological and evolutionary questions about host specificity, but if they are not carefully planned and carried out they can be dangerous. We believe that biologists should take a strong look at the ethics of manipulating host-ectoparasite associations in the field. Because many ectoparasites are vectors, or potential vectors of zoonotic diseases, the transference of ectoparasites among host species can have catastrophic consequences in natural ecosystems. ACKNOWLEDGMENTS To the Council for International Exchange of Scholars (CIES), the J William Fulbright Scholar Program, the interactive faculty of the Department of Ecology, Universidade do Estado do Rio de Janeiro, and Centro de Estudos Ambientais e Desen-volvimento Sustentável (Ceads/Uerj) for bringing the authors together for a course on vertebrate ectoparasites, and for providing the opportunity to carry out this research. CEL Esbérard received a CNPq grant (Process 152910/2004-0) and HG Bergallo received a productive grant. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocr |

{kind=link}
{kind=link}
{kind=link}