 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 8, December 2005, pp. 841-845 Detection of Rickettsia rickettsii in the tick Amblyomma cajennense in a new Brazilian spotted fever-endemic area in the state of Minas Gerais Elizângela Guedes, Romário C Leite, Márcia CA Prata*, Richard C Pacheco**, David H Walker***, Marcelo B Labruna**/+ Departamento de
Medicina Veterinária Preventiva, Escola de Veterinária, Universidade
Federal de Minas Gerais, Belo Horizonte, MG, Brasil *Embrapa Gado de Leite,
Juiz de Fora, MG, Brasil **Departamento de Medicina Veterinária Preventiva
e Saúde Animal, Faculdade de Medicina Veterinária e Zootecnia,
Universidade de São Paulo, Av. Prof. Orlando Marques de Paiva 87,
Cidade Universitária, 05508-270 São Paulo, SP, Brasil ***Department
of Pathology, University of Texas Medical Branch, Galveston, Texas, US Financial support: Fapesp (grant 03/13872-4 to MBL), Fogarty International Center (grant D43TW00903 to DHW), CNPq fellowship to MBL Received 4 August
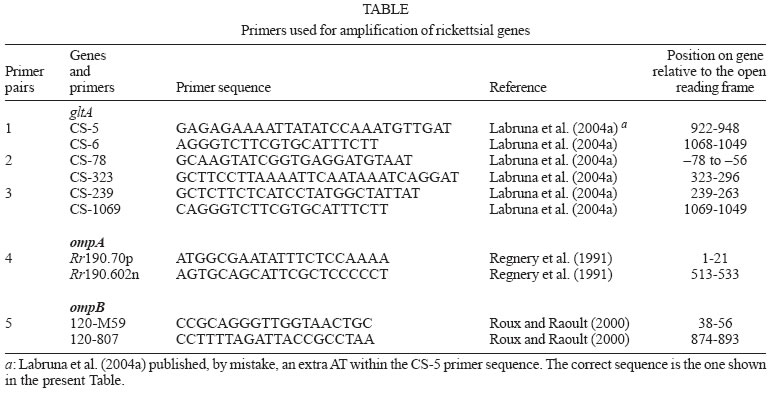
2005 Code Number: oc05172 The present study evaluated rickettsial infection in Amblyomma spp. ticks collected in a farm in Coronel Pacheco, a Brazilian spotted fever (BSF) endemic area. A total of 78 A. cajennense and 78 A. dubitatum free-living adult ticks were collected and tested by polymerase chain reaction (PCR) targeting a fragment of the rickettsial gene gltA. Only one pool of three A. cajennense ticks showed the expected product by PCR. This pool was further tested by PCR using sets of primers targeting the rickettsial genes gltA, ompA, and ompB. All reactions yielded the expected bands that by sequencing, showed 100% identity to the corresponding sequences of the Rickettsia rickettsii gene fragments gltA (1063-bp), ompA (457-bp), and ompB (720-bp). The minimal infection rate of R. rickettii in the A. cajennense population was 1.28% (at least one infected tick within 78 ticks).The present study showed molecular evidence for the presence of R. rickettsii in A. cajennense from a BSF-endemic area in Coronel Pacheco, state of Minas Gerais. Although R. rickettsii has been previously reported infecting A. cajennense ticks in Brazil and other Latin American countries, the present study performed the first molecular characterization of R. rickettsii from the tick A. cajennense. Key words: Amblyomma cajennense - Amblyomma dubitatum - Rickettsia rickettsii - Brazilian spotted fever Brazilian spotted fever (BSF) is a highly lethal rickettsial disease that has been reported in the Southeastern region of Brazil since the 1920s (Galvão et al. 2003). The disease is caused by the bacterium Rickettsia rickettsii, which is transmitted to humans through the bite of infected ticks. This bacterium has also been reported in the United States, Mexico, Costa Rica, Panama, and Colombia (Patino et al. 1937, Bustamante & Varela 1947, Rodaniche 1953, Philip et al. 1978, Fuentes 1979). In each of these countries, the disease in humans has received a different local designation (e.g., Rocky Mountain spotted fever in the United States, fiebre manchada in Mexico, fiebre de Tobia in Colombia). The tick species A. cajennense is the main vector of BSF. This statement is supported by several isolations of R. rickettsii from this tick species in the states of Minas Gerais (Moreira & Magalhães 1935, Dias et al. 1937) and São Paulo (Valejo-Freire 1946) and several epidemiological associations of the occurrence of BSF with high infestations by A. cajennense (Lemos et al. 1997, Horta et al. 2004a, Sangioni et al. 2005). In addition, several laboratory experiments have demonstrated that this tick species is an efficient experimental vector of R. rickettsii, which survives by transstadial and transovarial transmission in A. cajennense (Brumpt 1933, Parker et al. 1933, Travassos & Vallejo-Freire 1942). Besides Brazil, R. rickettsii has been isolated from A. cajennense ticks from Mexico, Pa-nama, and Colombia (Patino-Camargo 1941, Bustamante & Varela 1946, Rodaniche 1953). Other tick species incriminated as vector of R. rickettsii to humans are A. aureolatum in São Paulo, Brazil (Pinter & Labruna 2006), Rhipicephalus sanguineus in Mexico (Bustamante & Varela 1947) and Dermacentor andersoni and D. variabilis in the United States (Burgdorfer 1988). BSF has shown considerable reemergence since the 1980s. Many areas where the disease was never reported before are now considered endemic areas (Silva & Galvão 2004). One of these areas is Coronel Pacheco, within the administrative district of Juiz de Fora, Minas Gerais. From 2001 to 2004, at least 65 cases of BSF (11 were fatal; lethality: 17%) were reported in this district (Costa 2004). In addition, other seven lethal BSF cases were documented in 2004 in this area. In this context, the present study evaluated rickettsial infection in Amblyomma spp. ticks collected on a farm in Coronel Pacheco, a BSF endemic area. In addition, we provide the first molecular characterization of R. rickettsii detected in the tick A. cajennense. MATERIALS AND METHODS Study site and ticks - Free-living ticks were collected from a livestock pasture in the experimental farm of the Empresa Brasileira de Pesquisa Agropecuária – Centro Nacional de Pesquisa em Gado de Leite (Embrapa CNPGL), in Coronel Pacheco County (21°35' S, 43°15' W), state of Minas Gerais, Brazil. The area is 435 m above sea level. The weather is characterized by a warm rainy season from October through March and a dry season from April to September. Mean temperature ranges from 16°C in the winter to ≥ 23°C in the summer (Antunes 1986). The pasture was continuously inhabited by horses and dairy cattle. Domestic dogs were occasionally seen roaming in the pasture. In the lowland of the pasture, there was a small river that formed small marshes on both margins, which were inhabited by free-living capybaras that used to enter and cross the pasture to forage on corn crops located surrounding the pasture. This area was selected for the study because a recent, laboratory-confirmed case of BSF occurred in a human resident, who was infested by ticks after walking in this pasture a few days before onset of the disease. Free-living ticks were collected on the pastures from November 2003 to May 2004, on nine different dates at 4-35 day-intervals. On each date, four CO2 traps were mounted on the pasture, as described elsewhere (Oliveira et al. 2000). Collected ticks were brought alive to the laboratory where they were identified to species and incubated at 35°C and relative humidity ≥ 80% for three to four days to increase the rickettsial burden (Burgdorfer 1970). Thereafter, they were processed by the hemolymph test as described below. Hemolymph test - Ticks were individually processed by the hemolymph test as described (Burgdorfer 1970). Briefly, a drop of hemolymph of each tick was dried on a glass slide and stained by the Giménez method (Giménez 1964). Thereafter, ticks were frozen at -20oC until processed for molecular analyses. Molecular analyses of ticks - Ticks were processed in pools of three specimens of the same species. Each group of three frozen ticks was thawed in a water bath at 37oC, and pooled in a sterile 1.5-ml microtube containing sterile phosphate-buffered saline (PBS), where they were cut into small pieces with a sterile scissors and homogenized with a sterile micropestle. A volume of 200 µl of tick homogenate was aspirated through a 21-gauge needle attached to a 1-ml syringe, and DNA was extracted using the Dneasy Tissue Kit (Qiagen, Chatsworth, CA) following the manufacturer's protocol for isolation of DNA from animal blood samples. Previous studies have shown that this protocol of DNA extraction was successful for releasing rickettsial DNA in tick tissues (Labruna et al. 2004a, b). Purified DNA was stored at 4oC until used as template for polymerase chain reaction (PCR) amplifications. Five µl of template (approximately 500 ng of tick DNA) were used for each PCR. Each tick pool was processed by PCR using the primers CS-5 (forward) and CS-6 (reverse) (Table) that amplify a 147 bp fragment of the citrate synthase gene (gltA) of spotted fever group rickettsiae. These primers have been shown to have a sensitivity down to a single copy of R. rickettsii DNA (Labruna et al. 2004a). Once a tick pool was positive by PCR, it was tested in a battery of PCRs using the remaining primer pairs shown in Table, targeting fragments of the rickettsial genes gltA, ompA and ompB, as described elsewhere (Labruna et al. 2004a,b). All PCR reactions (50 µl final volume) were performed by adding 5 µl of DNA template to 25 µl of the PCR iQTM Supermix (BioRad), 1 µl (20 pmoles) of each primer, and 18 µl of molecular grade water. PCR cycling conditions for primers CS-5 and CS-6 were 1 cycle at 95oC for 2 min followed by 50 cycles of 15 s at 95oC, 30 s at 50oC, and 30 s at 60oC. For the other primer pairs, PCR cycling conditions were 1 cycle at 95oC for 3 min, 40 cycles of 15 s at 95oC, 30 s at 50oC, 30 s at 72oC, followed by 1 cycle at 72oC for 7 min. For each reaction, three negative controls (5 µl of the same water used above) and a positive control [300 ng of DNA of R. parkeri-infected A. cajennense tick (Sangioni et al. 2005)] were included. Ten microliters of each PCR product was separated by electrophoresis in a 2% agarose gel, stained with ethidium bromide and examined using UV transillumination. PCR products of the expected size were purified using ExoSap (USB) and sequenced in an automatic sequencer (Applied Biosystens/Perkin Elmer, model ABI Prism 310 Genetic, California, US) according to the manufacturer's protocol. Partial sequences obtained were submitted to BLAST analysis (Altschul et al. 1990) to determine similarities to other Rickettsia species. RESULTS During field collection of ticks, a total of 78 (17 males and 61 females) A. cajennense and 78 (31 males and 47 females) A. dubitatum adult ticks were collected. No tick specimen demonstrated Rickettsia-like organisms by the hemolymph test. Some ticks (131) yielded hemolymph containing none or just a few cells whereas hemolymph cells from other ticks (25) were lost during the staining process through the washes. By PCR, one pool of three A. cajennense female ticks yielded the expected product with the primers CS-5 and CS-6. The three ticks that composed this pool had their hemolymph lost during the hemolymph cells test. When this same tick pool was tested by PCR with the other primer pairs described in Table, all reactions yielded the expected bands. When the amplified fragments were sequenced, the gltA fragment showed to be 100% (1063/1063) similar to the corresponding sequence (DQ115890) of R. rickettsii isolated from the tick A. aureolatum in the state of São Paulo, and 99.7% (1060/1063) similar to the sequence (U59729) of the Bitterroot strain of R. rickettsii isolated from the tick D. andersoni in the United States. The ompA fragment was 100% (457/457) similar to the corresponding sequence (U43804) of the Bitterroot strain of R. rickettsii, and 99.6% (455/457) similar to the sequence (AY319293) of the Hlp#2 strain of R. rickettsii isolated from the tick Haemaphysalis leporispalustris in the United States. The ompB fragment was 100% (720/720) similar to the corresponding sequence (X16353) of the Bitterroot strain of R. rickettsii. DISCUSSION The present study provided molecular evidence for the presence of R. rickettsii in at least one A. cajennense tick collected in a pasture of a BSF-endemic area in Coronel Pacheco, state of Minas Gerais. Although R. rickettsii has been reported infecting A. cajennense ticks in Brazil and other Latin American countries (Moreira & Magalhães 1935, Dias et al. 1937, Patino-Camargo 1941, Bustamante & Varela 1946, Valejo-Freire 1946, Rodaniche 1953), the present study reports the first molecular identification of R. rickettsii in A. cajennense ticks. The DNA sequence of the rickettsial gltA gene generated in the present study showed to be identical to the corresponding sequence of another Brazilian R. rickettsii strain, isolated from A. aureolatum (Pinter & Labruna 2006). These two sequences were 99.7% (1060/1063) similar to the Bitterroot strain from the United States. This slight polymorphism was due to an insertion or deletion (indel) of the codon CCG at the position 865-867 of the ORF of the R. rickettsii gltA gene (Pinter & Labruna 2006). Interestingly, Eremeeva et al. (2003) reported that three Latin American strains of R. rickettsii (Brazil, Colombia, and Costa Rica) and some from United States had this specific codon on their gltA sequence whereas other North American strains lacked it. The origin or significance of such polymorphism remains unknown. The ompA partial sequence of R. rickettsii generated in the present study showed to be identical to the corresponding sequence of the Bitterroot strain from the United States, but slightly different (99.6%) from another R. rickettsii strain (Hlp#2) isolated from the tick H. leporispalustris in that same country. Hlp#2 is a less virulent strain, which has never been associated with human disease. In contrast, the Bitterroot strain is considered to be highly virulent (Eremeeva et al. 2003). Thus, the R. rickettsii genotype detected in the tick A. cajennense of the present study possibly corresponds to the one related to the fatal cases of BSF in the Coronel Pacheco endemic area We tested 26 pools of A. cajennense and 26 pools of A. dubitatum, each pool containing three ticks. As only one pool of A. cajennense was shown to contain rickettsial DNA, we determined that the minimal infection rate of R. rickettsii in the A. cajennense population of that pasture was 1.28% (at least one infected tick within 78 ticks). This value is remarkably high if compared to previous studies performed on A. cajennense and other tick species in Brazil and United States. In the state of São Paulo, Sangioni et al. (2005) tested by PCR 810 A. cajennense specimens from three BSF-endemic areas, but no tick yielded rickettsial DNA. In another BSF endemic area in the state of São Paulo, Pinter and Labruna (2005) detected R. rickettsii in six (0.89%) out of 669 A. aureolatum ticks. In the United States, the infection rate of R. rickettsii reported among D. variabilis populations from Rocky Mountain spotted fever endemic areas has varied from 0.05 to 1.3% (Burgdorfer 1988). Our finding of at least a 1.28% infection rate of A. cajennense ticks by R. rickettsii could not represent the real scenario in the field, due to our small tested sample (78 tick specimens). On the other hand, this result is in accordance with the high number of BSF cases (65) reported in this region during the last five years (Costa 2004). In the present study, none of the A. dubitatum ticks contained rickettsial DNA. This result contrasts with other studies that reported A. dubitatum (formerly A. cooperi) to be commonly infected by other Rickettsia species, such as R. bellii (Labruna et al. 2004a), and less commonly, by a rickettsia strain closely related to Rickettsia parkeri (Labruna at al. 2004a). Our study comfirms that R. rickettsii is circulating in A. cajennense ticks from the BSF-endemic area of Coronel Pacheco. Although we found a calculated high infection rate of this agent among its vector population, further long-term studies should evaluate the dynamics of the R. rickettsii infection rate in A. cajennense populations. Such studies have never been performed, but should provide important information for understanding the irregular pattern and the currently unpredictable occurrence of BSF in endemic areas. Finally, these studies should also take in account the rickettsial infection in other tick species, such as A. dubitatum, and the faunal composition of the endemic area. For instance, capybaras (present in large numbers in the area of the present study) are a primary host for A. cajennense and have long been suspected to be a competent amplifier host of R. rickettsii among Amblyomma ticks (Travassos & Vallejo-Freire 1942, Lemos et al. 1996, Souza et al. 2004). ACKNOWLEDGMENTS To John Furlong for the valuable support during experiment, to Embrapa Gado de Leite for technical support during field work. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05172t1.jpg] |
| |||||||||

{kind=link}