 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 8, December 2005, pp. 875-881 Clinical and pathological importance of vacA allele heterogeneity and cagA status in peptic ulcer disease in patients from North Brazil Luisa Caricio Martins/+, Tereza Cristina de Oliveira Corvelo, Samia Demachki*, Marialva TF Araujo*, Mônica Baraúna Assumpção**, Simone Cristina Araujo Jucá Vilar***/****, Felipe Bonfim Freitas, Hivana Patricia Melo Barbosa, Amanda Alves Fecury, Renata Kelly Costa do Amaral, Sidney Emanuel Batista dos Santos***** Laboratório
de Imunogenética, Departamento de Genética, Centro de Ciências
Biológicas, Universidade Federal do Pará, Rua Augusto Correa
s/no, Campus Universitário do Guamá, 66075-110
Belém, PA, Brasil *Departamento de Patologia, **Departamento de Endoscopia,
Hospital Universitário João de Barros Barreto Belém,
PA, Brasil ***Departamento de Endoscopia, Hospital Ofir Loiola, Belém,
PA, Brasil ****Departamento de Endoscopia, Hospital Universitário
Betina de Ferro Souza, Belém, PA, Brasil *****Laboratório de
Genética Humana e Médica, Departamento de Patologia, Universidade
Federal do Pará, Belém, PA, Brasil Financial support: Sectam, Capes Received 13 July
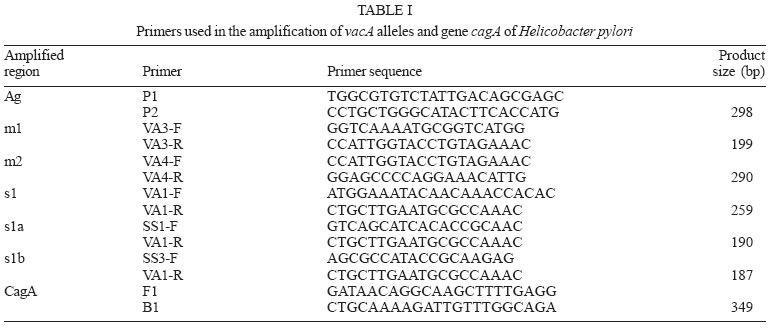
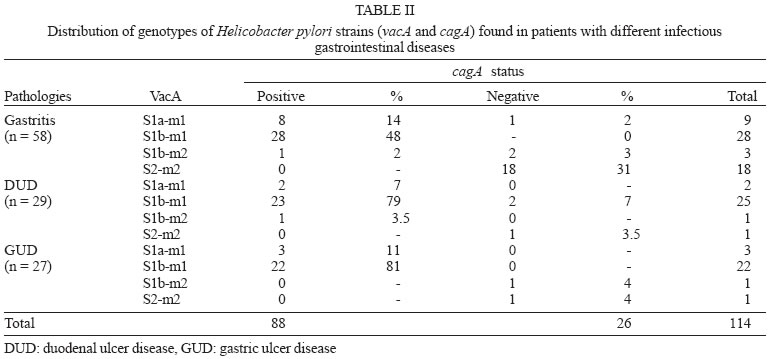
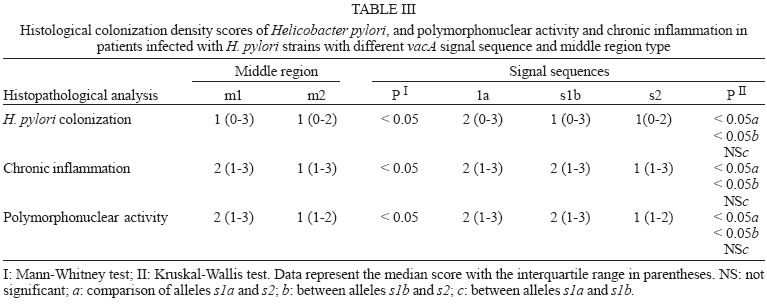
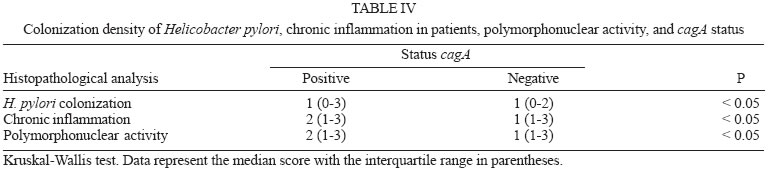
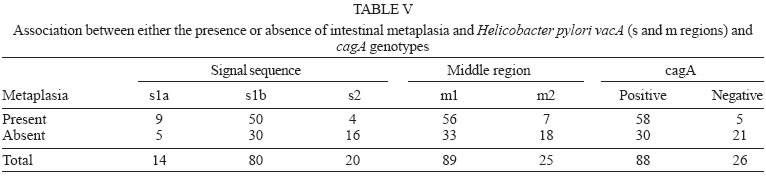
2005 Code Number: oc05177 We have examined the prevalence of gene cagA and vacA alleles in 129 patients, 69 with gastritis and 60 with peptic ulcer diseases from North Brazil and their relation with histopathological data. vacA and cagA genotype were determined by polymerase chain reaction. Hematoxylin-eosin staining was used for histological diagnosis. 96.6% of the patients were colonized by Helicobacter pylori strains harboring single vacA genotype (nont-mixed infection). Among them, 11.8% had subtype s1a, 67.8% had subtype s1b, and 17% subtype s2. In regard to the middle region analysis, m1 alleles were found in 75.4% and m2 in 21.2% of patients. The cagA gene was detected in 78% patients infected with H. pylori and was associated with the s1-m1 vacA genotype. The H. pylori strains, vacA s1b m1/cagA-positive, were associated with increased risk of peptic ulcer disease and higher amounts of lymphocytic and neutrophilic infiltrates and the presence of intestinal metaplasia. These findings show that cagA and vacA genotyping may have clinical relevance in Brazil. Key words: Helicobacter pylori - peptic ulcer - vacA alleles - cagA status - Brazil Helicobacter pylori infection is the major cause of chronic superficial gastritis in humans, an important etiological factor in the pathogenesis of peptic ulcers (PUD) and some forms of gastric cancer (Dunn et al. 1997). However, most people who harbor H. pylori are asymptomatic, and only a few patients infected with this bacterium develop either PUD or gastric cancer (Nomura et al. 1994). The variability of clinical manifestations is associated with several factors such as specific organism virulence factor, host immunological factor, and environmental influences, or their combination (Taylor & Blaser 1991, Atherton et al. 1997). Cytotoxin VacA and protein CagA are the two major virulence markers usually associated with H. pylori pathogenicity. The vacuolating cytotoxin (VacA) induces the formation of intracellular vacuoles in epithelial cell lines (Cover & Blaser 1992, Atherton et al. 1995). Besides its direct cell-damaging effect in vitro, VacA also plays a major role in inducing cytoskeletal changes, apoptosis, suppression of epithelial proliferation, and migration (Pai et al. 1999, Cover et al. 2003). vacA gene is present in all H. pylori strains and contains at least two variable parts. The s-region, which encodes the signal peptide, coexists as either s1 or s2 allelic types. Subtypes s1a, s1b, and s1c have been identified among type s1 strains. The m-region (middle) occurs as m1 or m2 allelic types (Atherton et al. 1995, Papini et al. 2001). Production of VacA is related to the mosaic structure of vacA. In general, type s1/m1 and s1/m2 strains produce high and moderate levels of toxin, respectively, whereas s2/m2 strains produce little or no toxin (Atherton et al. 1995, 1997, Forsyth et al. 1998). Recent in vitro work has demonstrated that strains with genotype vacA s1/m1, but without s2/m2 mediate channel activity and cellular vacuolation in intoxicated cells, which results in a detectable reduction of mitochondrial transmembrane potential and the release of cytochrome c. This suggests a functional correlation between VacA channel activity and changes in mitochondrion membrane permeability (Willhite & Blanke 2004). A second disease-associated virulence factor is the cytotoxin-associated protein (CagA), a high molecular mass immunodominant protein (140 KDa) that is part of the cag pathogenicity island (PAI) (Tummuru et al. 1995, Segal et al. 1999). Gene cagA is considered a marker for the presence of Cag PAI, which is associated with the most virulent H. pylori strains. Injected into the cytoplasm of the host cell, protein CagA plays an important role in the induction of cellular signaling and elicits multiple significant morphological changes, proliferation, and apoptosis (Stein et al. 2000, Hatakeyama 2003). Studies conducted in several countries have shown that vacA-type s1 and cagA-positive H. pylori strains are associated with severe H. pylori-induced PUD (Atherton et al. 1995, van Doorn et al. 1998, Rudi et al. 2000, Catalano et al. 2001). Recent publications have also uncovered this association in Brazil (Mattar & Sandanna 2000, Brito et al. 2000, Ashour et al. 2002). In contrast, many reports from several parts of the world (Pan et al. 1997, 1999, Zheng et al. 2000, Backert et al. 2004) do not describe any significant association between genotypes (vacA type s1, cagA positive) and clinical responses among the various groups of infected patients with chronic gastritis, ulcer, and gastric cancer. Thus, existing data is contradictory and cannot explain the pathogenic role of H. pylori in the development of different gastric diseases. Consequently, it might be useful to know the genetic diversity of H. pylori strains in North Brazil, a region with a high frequency of H. pylori-induced gastric disease. The aim of this study was to determine the prevalence of gene cagA and vacA alleles in adult patients with gastritis and PUD in our population as well as the relation between cagA and/or vacA genotypes and histopathological data in the development of PUD. MATERIALS AND METHODS This study was approved by the Ethics Committee of Hospital Universitário João de Barros Barreto, Belém, PA, Brazil. All patients gave their informed consent to participate in the experiment. Patients - This study was carried out at Hospital Universitário João de Barros Barreto, Hospital Univer-sitário Betina de Ferro Souza, and Hospital Ofir Loiola from September 2002 to January 2003. During this period, gastric biopsies of 129 consecutive patients (median age, 44 years; range, 15-75 years; 60 women and 69 men) who underwent gastric endoscopy due to different digestive disorders were obtained. None of the patients had received antimicrobial drugs, H2-receptor antagonists, acid pump inhibitors, nonsteroidal anti-inflammatory drugs, or any medication for at least 60 days before endoscopy. During endoscopy, six biopsy fragments were taken from the stomach of each patient. Two antral and two corpus gastric biopsies were analyzed by histological methods, and two antrum specimens were also analyzed by molecular methods. Histological evaluation - The biopsy specimens were fixed in 10% buffered formalin solution, embedded in paraffin, cut into sequential 0.4-µm sections, and stained with hematoxylin and eosin (H&E). The histopathological parameters were graded 0-3 using the criteria described in the updated Sydney classification system (Dixon et al. 1997) for analysis of chronic inflammation, polymorphonuclear activity, and bacterial density. Whenever present, intestinal metaplasia was typed as either complete, or mixed (complete and incomplete), or incomplete. The presence of H. pylori in the sections was determined by using a modified Gram staining protocol and taking into consideration its morphological characteristics: curved and spiral form and intense blue colouring. DNA isolation - Total DNA was extracted from frozen gastric biopsy specimens using the following procedure: 10 µl of proteinase K and 300 µl of lysis buffer (200 mM Tris-HCl, 25 mM EDTA, 300 mM NaCl, 1.2% sodium dodecyl sulfate) were added to the biopsy specimens. The mixture was incubated at 55°C for 12 h. The lysate was extracted with an equal volume of phenol-chloroform, precipitated with isopropanol, and washed with 70% ethanol. The pellet was dried and suspended in 200 ml of sterile distilled water. DNA extracts were stored at -20°C. PCR amplification and detection of amplified DNA products - One set of primers (p1 and p2) (Hammar et al. 1992) that amplifies a gene fragment of 298 bp present in all strains of H. pylori was used to detect bacterial DNA. Only positive samples were used for further study. Amplification of vacA signal sequences and middle regions was performed by PCR with oligonucleotide primers described by Atherton et al. (1995). The strains were initially characterized as either type s1 or s2 and as either type m1 or m2. All s1 strain alleles were further characterized as either variant s1a or s1b. The previously described F1 and B1 primers were used (Tummuru et al. 1993) to detect CagA as shown in Table I. All PCR mixtures were prepared in a volume of 25 µl containing 0.5 nM of each primer; 1X PCR buffer; 1.5 nM MgCl2; sterilized water, 0.2 nM deoxynucleoside; 1.25µl Taq DNA polymerase, and 2 µl DNA sample. The mixtures were placed in a thermal cycler. PCR amplification was performed under the following conditions: initial denaturation at 95°C for 2 min followed by 35 cycles of denaturation at 95°C for 1 min, annealing and extension for 1 min, and final extension at 72°C for 10 min. Annealing temperatures were set at 58°C for primers VA3-F/VA3-R, VA4-F/VA4-R, F1/B1, and at 63°C for SS1-F, SS3-F, SS2-F/ VA1-R. Negative and positive controls were used in all reactions. PCR products were visualized by electrophoresis in 2% agarose gel, stained with ethidium bromide, and examined under UV illumination. Statistical evaluation - Data were analyzed with Biostat version 3.0 software (Ayres et al. 2003). Chi-square tests, G test, and odds ratio (OR) with 95% confidence intervals (95% CI) were used to assess the association between vacA alleles and cagA gene and the development of PUD. Histopathological parameters were analyzed by the Kruskal-Wallis test and Mann-Whitney test. Differences were considered statistically significant for P values less than 0.05. RESULTS Distribution of vacA alleles and cagA gene - Genotype vacA was detected in the antral mucosa of 91.5% (118/129) of the patients studied. 96.6% (114/118) of patients were colonized by H. pylori strains harboring single vacA genotype (non-mixed infection). Among them, the s1 signal sequence was detected in 79.6% (94/114), of which 11.8% (14/114) had subtype s1a, and 67.8% (80/114) had subtype s1b. Subtype s2 was detected in 17% (20/114) patients. With regard to analysis of the middle region, alleles m1 and m2 were found in 75.4% (89/114) and 21.2% (25/114) patients, respectively. Three combinations of signal sequences and middle region types were also identified. Among the 114 patients with non-mixed infection, strains with genotype s1a/m1 were found in 12.3% (14/114) patients, genotype with s1b-m1 in 65.7% (75/114), and with genotype s1b/m2 in 4.4% (5/114) patients. Genotype s2/m2 was detected in 17.6% (20/114) patients. No strains presented genotype s2/m1. Multiple strain infection (colonization of the same patients by H. pylori strains harboring more than one vacA genotype) was detected in 3.4% (4/118) patients. Strains with s1b/m1 - s2/m2 genotype were detected in 2.6% (3/118) patients and with s1a/m1 - s2/m2 in 0.8% (1/118) patient. None of the patients with multiple strain infection were included in the analysis of the comparison of vacA genotype and cagA gene or their relationship to gastrointestinal diseases. Gene cagA was detected in 92 out of 118 (78%) patients infected with H. pylori; 75% (88/114) had no mixed infection. The presence of gene cagA was strongly associated with the s1-m1 vacA genotype (G = 476.7598, p < 0.001). Relation between H. pylori genotypes and PUD - Among the 129 patients selected, 69 (53.5%) had gastritis, 29 (22.5%) had gastric ulcer disease (GUD), and 31 (24%) had duodenal ulcer disease (DUD). Seven of the patients with gastritis did not present H. pylori DNA, and four were colonized by two or more H. pylori strains. Two patients with GUD and two with DUD were H. pylori negative and none presented mixed infection. The genotypes of H. pylori strains found in patients with different gastrointestinal diseases are given in Table II. Genotype s1/m1/cagA-positive is the most frequent among patients with DUD (p = 0.01, OR = 7.09, 95% CI = 1.52-32.48) and GUD (p = 0.00, OR = 7.09, 95% CI = 1.52-32.98). However, a separate analysis of the frequency of each allele in the different pathologies revealed an association between allele s1b and the presence of DUD (p = 0.00, OR = 15.09, 95% CI = 1.88-120.85) and GUD (p = 0.00, OR = 13.35, 95% CI = 1.66-107.40). In this study, allele s1a was not statistically associated with the occurrence of either DUD (p = 0.61, OR = 4.00, 95% CI = 0.31-50.23) or GUD (p = 0.29, OR = 6.00, 95% CI = 0.54-66.17). In relation to alleles m1 and m2, it was observed that the presence of allele m1 was associated with increased risk of DUD (p = 0.00, OR = 7.66, 95% CI = 1.65-35.48) and GUD (p = 0.00, OR = 7.09, 95% CI = 1.52-32.98). Gene cagA also displayed an association with the development of DUD (p = 0.02, OR = 4.91, 95% CI = 1.32-18.22) and GUD (p = 0.00, OR = 7.09, 95% CI = 1.52-32.98). Relation between H. pylori genotypes and histological evaluation - All 114 patients who were infected by only one strain of H. pylori presented chronic gastritis in the antral region. However, the bacterium was observed in the gastric tissue of 80 (70%) of the patients by the histologic method. Comparing the degree of colonization, inflammation, and neutrophilic activity and vacA alleles of infecting strains, it was observed that patients infected with alleles s1 and m1 presented a degree of colonization, inflammation, and neutrophilic activity larger than that of allele s2 and m2 (Table III). However, the comparison of histological scores (colonization density, degree of inflammation, and neutrophilic activity) observed in patients infected by strains with subtypes allele s1a and those infected by s1b did not reveal a statistically significant difference (Table III). In relation to cagA positive strains, higher levels of colonization, inflammation, and neutrophilic activity were also verified in patients who were infected with strains of cagA-negative (Table IV). Of the 114 patients infected by only one strain of H. pylori, 63 (55.3%) presented incomplete intestinal metaplasia. The risk of developing metaplasia was larger in patients infected by strains that had allele s1a (p = 0.02, OR = 7.20, 95% CI = 1.53-33.84), s1b (p = 0.00, OR = 6.66, 95% CI = 2.03-21.81), m1 (p = 0.00, OR = 4.36, 95% CI = 1.64-11.54) and cagA-positive (p = 0.00, OR = 8.12, 95% CI = 2.78-23.68) when compared with strains with alleles s2, m2, and cagA-negative, respectively (Table V). DISCUSSION Infection by H. pylori is cosmopolitan and one of the most frequent in the whole world. The pathogen, a genetically diverse species with a high DNA recombination rate, may be involved in the complex variety of diseases in infected patients (Nomura et al. 1994, Dunn et al. 1997). Studies carried out in several countries have demonstrated differences in the distribution of vacA alleles and the presence of cagA gene in genotypes of strains of H. pylori and their association with the development of gastroduodenal diseases (Pan et al. 1997, 1999, Kidd et al. 1999, Morales-Espinosa et al. 1999, Zheng et al. 2000, Backert et al. 2004). In this study we have found that 78% of the H. pylori species isolated from the patients had the vacA s1-m1 genotype, which is a higher value than described for other Brazilian cities, such as Belo Horizonte, MG, 35% (Ashour et al. 2002), Bragança Paulista, SP, 59% (Ribeiro et al. 2003), and Marília, SP, 59% (Gatti et al. 2005). This result indicates that the genotype frequency of vacA in Brazilian regions is different from one to the other. In the present study, evidence was found of the presence of multiple strains, based on vacA genotyping in 3.4% of the cases. Similar results were found by Ribeiro et al. (2003). Other studies of Brazilian patients found 13.4% (Ashour et al. 2001) and 11% (Gatti et al. 2005) coinfection by multiple strains. The prevalence of multiple H. pylori strains may be still underestimated, especially in areas with a high prevalence of H. pylori infection (van Doorn et al. 1998, Ashour et al. 2001). The sequencing of the s and m region of the vacA gene can define the degree of variation present and thus characterize the differences between the H. pylori species in the different regions. In this study, we observed that genotype s1b-m1/cagA-positive was the most frequent in patients with PUD. This variant is also more frequent in strains obtained from patients from Portugal, Spain, and African countries with historic, cultural and economic relationships with Brazil (Kidd et al. 1999, van Doorn et al. 1999, Noqueira et al. 2001). High prevalence of genotype s1b/m1/cagA-positive has also been detected in association with the development of PUD in north European patients (Nogueira et al. 2001, Tham et al. 2001). Our data indicate that individuals colonized with s1b-m1/cagA-positive H. pylori strains are at increased risk of developing peptic ulceration. The association of the different vacA alleles and cagA gene with the development of gastrointestinal diseases is contradictory (Pan et al. 1997, 1999, Zheng et al. 2000, Backert et al. 2004). Other Brazilian studies have also demonstrated an association between s1b-m1/cagA-positive and the presence of PUD (Evans et al. 1998, Ashour et al. 2002, Ribeiro et al. 2003, Gatti et al. 2005). However, another study carried out in Recife, PE, Northeast region of Brazil, did not correlate the predominance of allele s1b (90.7%) with the development of PUD (Brito et al. 2003). These authors observed an association between the high predominance of gene cagA and duodenal ulcer. Similarly to what was observed for gene vacA, divergences in relation to the association of gene cagA and the development of gastric diseases were also reported relative to the geographic distribution of gene cagA in patients with PUD, which varied between 80 and 100%. In Europe and North America, it has been demonstrated that infection by cagA-positive strains increases the risk of developing atrophic gastritis and gastric cancer; however, other studies carried out with European and Asian populations did not confirm these data (Maeda et al. 1998, Zheng et al. 2000, Backert et al. 2004, Zhou et al. 2004). Persistent infection by H. pylori leads to an infiltration of neutrophils and lymphocytes in the gastric epithelia, causing a severe inflammatory activity, which can result in atrophic gastritis with development of intestinal metaplasia and displasia (Nogueira et al. 2001). In our study, we also observed that alleles s1b, m1, and the presence of gene cagA presented high degrees of colonization, inflammation, and neutrophilic activity when compared to alleles s2, m2, and the absence of cagA. Atherton et al. (1997) reported the association of allele m1 and histological alterations as related to an increase in epithelial damage (epithelial degeneration, mucus depletion, and microscopic erosions) in contrast to reports on allele m2. Similarly, Nogueira et al. (2001) observed that alleles s1b, m1 and the presence of gene cagA are associated to high degrees of gastric tissue inflammation and neutrophilic activity in Portuguese patients in relation to alleles s2, m2, and the absence of cagA. The presence of intestinal metaplasia in the gastric mucosa was observed in 40% of the Portuguese and 60% of the Colombian population, in association with alleles s1b, m1, and the gene cagA (Nogueira et al. 2001). For patients with chronic gastritis from Belém, PA, the study by Aguiar et al. (2002) described 40% as having intestinal metaplasia and in our study 55% had the pre-malign lesion associated with H. pylori infection of the species containing alleles s1b, m1, and gene cagA. Based on our study and others previously reported for the Brazilian population, we conclude that the strain containing genotypes s1b, m1, cagA-positive is the most prevalent one in patients with PUD. Our observations confirm the association of genotype s1b/m1/cagA-positive of the infecting strain and the development of PUD. Thus, the detection of VacA and CagA, virulence markers described in several clinical outcomes may be used to help the treatment and prevention of H. pylori infection. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05177t4.jpg] [oc05177t1.jpg] [oc05177t3.jpg] [oc05177t5.jpg] [oc05177t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}