 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 8, December 2005, pp. 893-902 Effect of food on immature development, consumption rate, and relative growth rate of Toxorhynchites splendens (Diptera: Culicidae), a predator of container breeding mosquitoes D Dominic Amalraj+, N Sivagnaname, PK Das Vector Control
Research Centre, Pondicherry, India Received 22 June
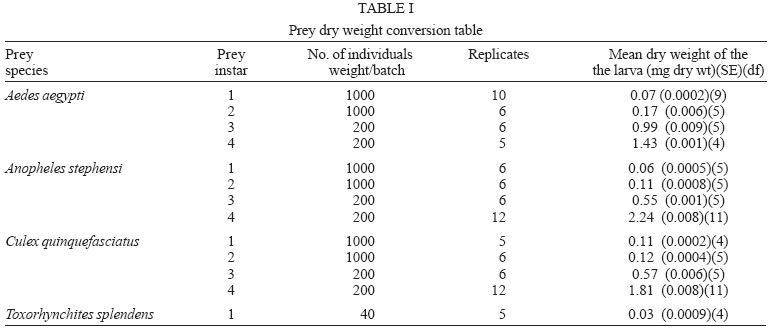
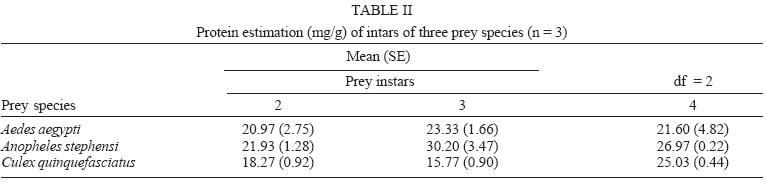
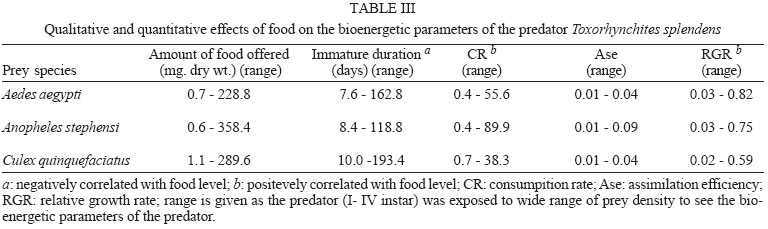
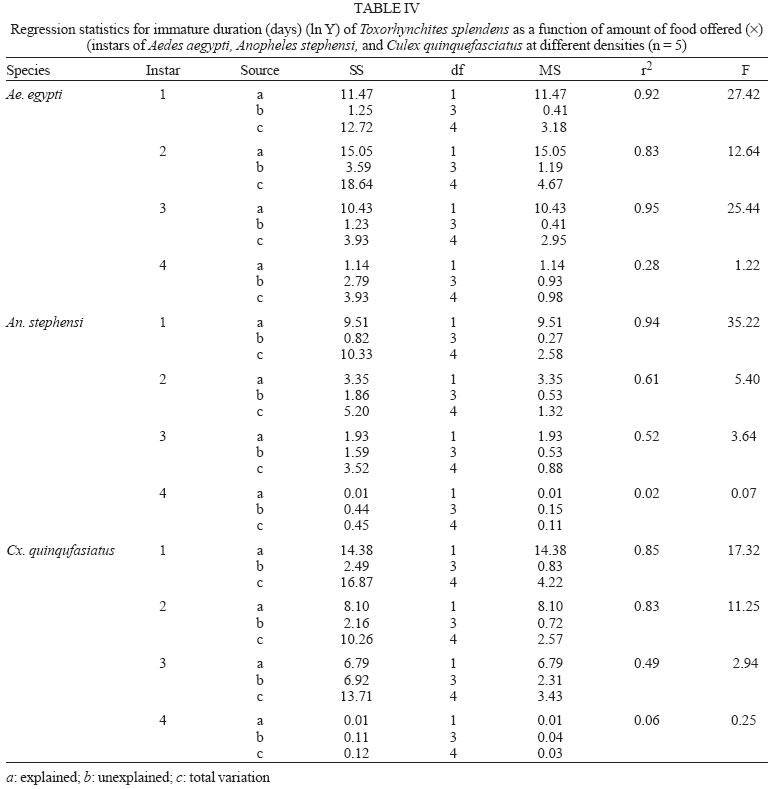
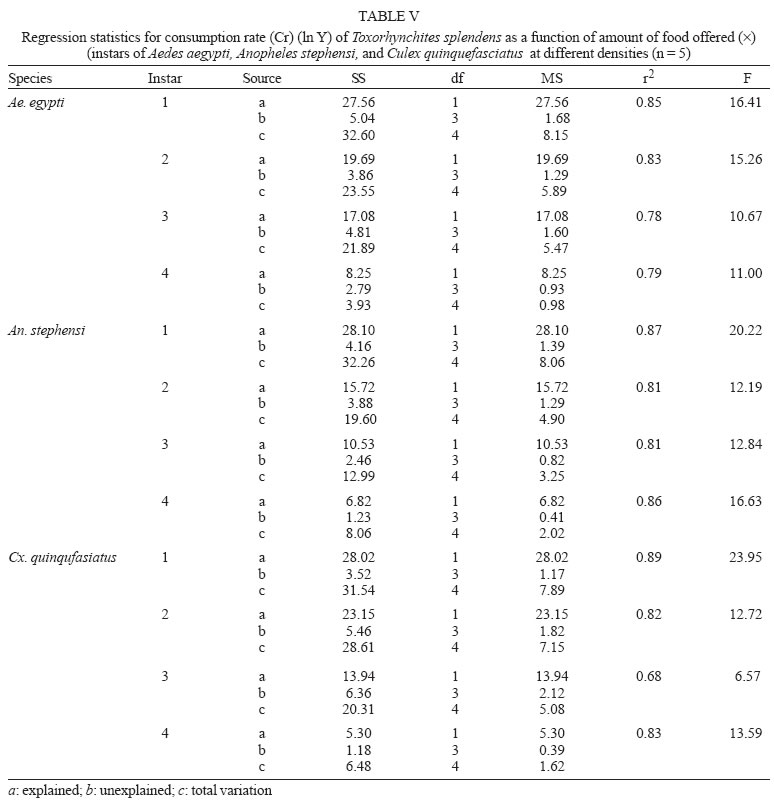
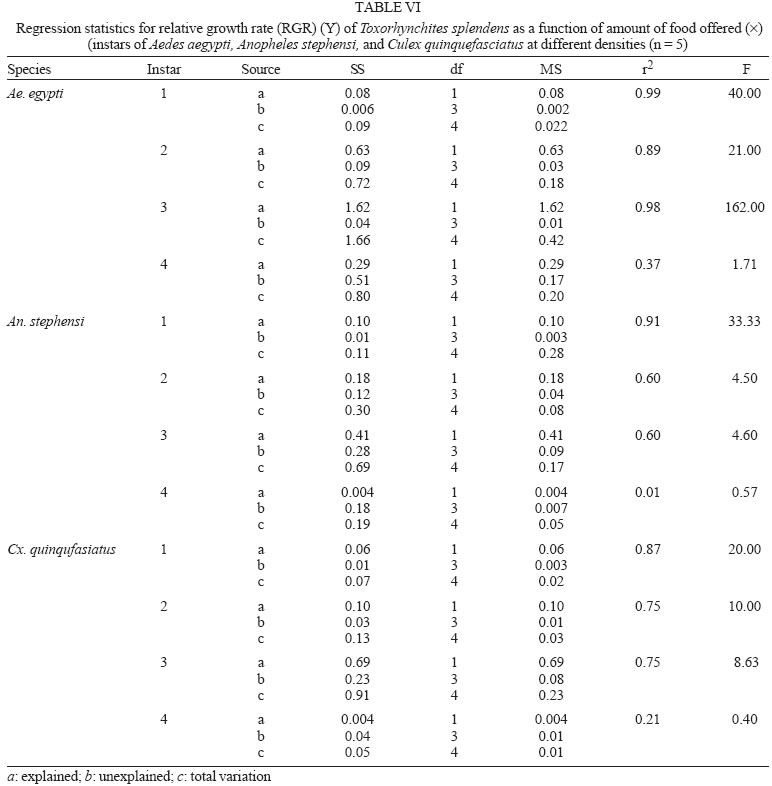
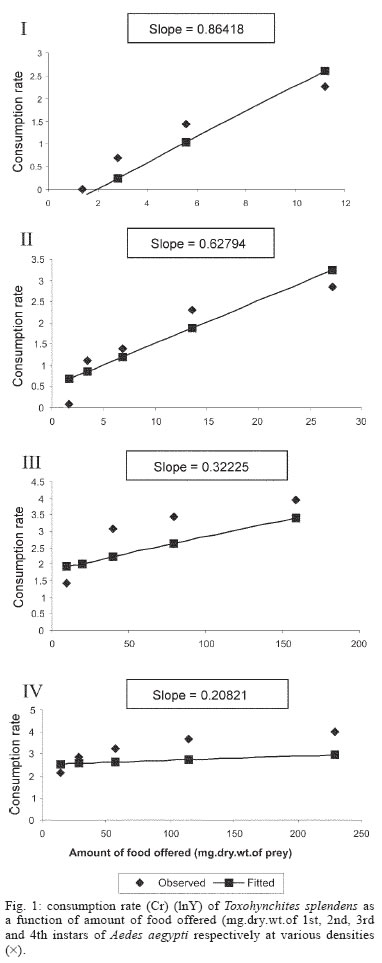
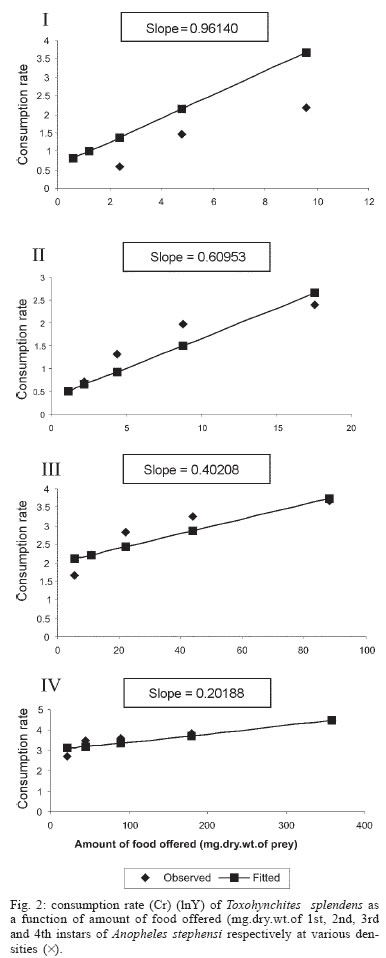
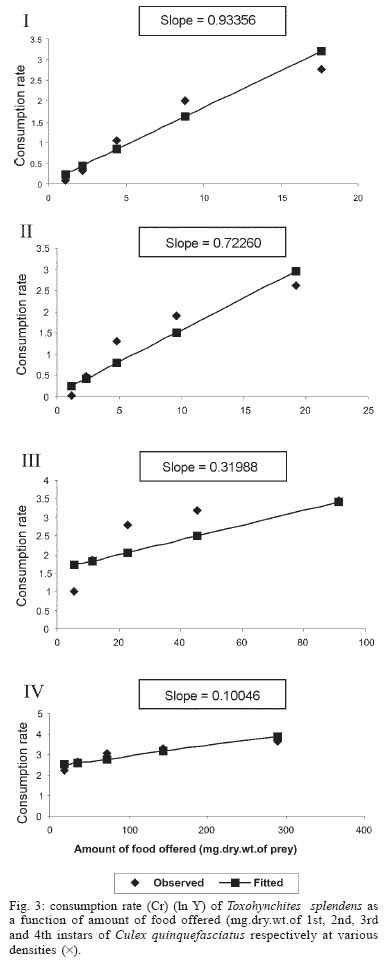
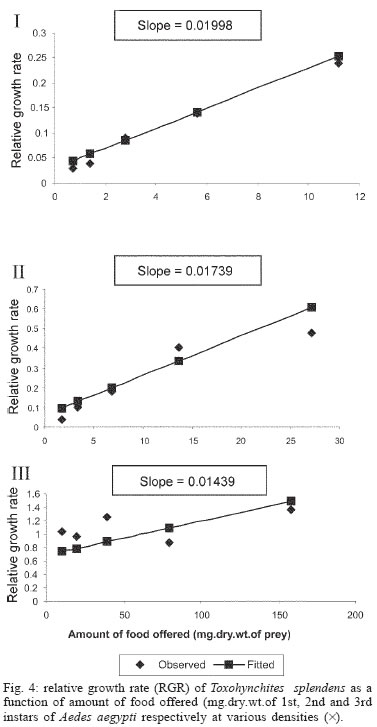
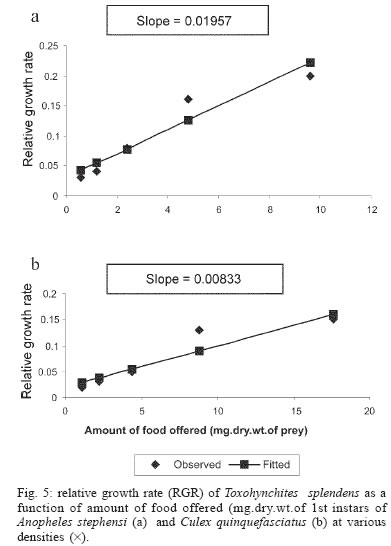
2005 Code Number: oc05180 Food utilization by the larvae of Toxorhynchites splendens (Wiedemann) was studied in the laboratory by offering larvae of Aedes aegypti Linnaeus, Anopheles stephensi (Liston), and Culex quinquefasciatus (Say). Quantitative analyses of data indicated that immature development was significantly faster with increase in food availability. The regression analysis showed that the degrees of the relationship between immature duration (Id) and food availability were higher when offered early instars of prey (first and second instars) than late instars. Consumption rate (Cr) of the predator increased with increase in food availability and this relationship was highly significant when larvae of An. stephensi were offered as food. Consumption rate to food level decreased with increase in the age class of the prey. There was a significant negative correlation between Id and Cr. This aspect helps to increase population turnover of T. splendens in a shorter period when the prey is abundant. Conversely, the predator compensated the loss in daily food intake at low food level by extending Id thereby attains the minimum threshold pupal weight for adult emergence. There was an increase in the relative growth rate (RGR) of the predator when An. stephensi was offered as prey and this was related to the high protein content of the prey per body weight. There was a positive correlation between Cr and RGR. This adaptive life characteristic strategy of this predator is useful for mass-rearing for large scale field release programmes in the control of container breeding mosquitoes is discussed. Key words: mosquito prey - predator - immature duration - consumption rate - relative growth rate - Toxorhynchites splendens A self-sustained biological control method is yet to be devised for mosquitoes. All available methods, including larvivorous fish, require repeated releases or treatments. Members of the genus Toxorhynchites were released in several Pacific islands to control container- breeding mosquitoes such as Aedes aegypti Linnaeus and Aedes albopictus (Garcia 1982). All these studies indicated that both larvae and adults of the predator should be released in large numbers to overcome the problem of low egg production and low progeny found in the field condition. Further, the low egg production in this genus in spite of the provision of adequate larval food poses a major drawback in mass production for field releases. Blumberg and Swiraki (1974) pointed out the influence of quality of food on food selection and feeding rate. Such differences in the quality of food may also have considerable influence on growth and survival of insects. Growth and development of T. rutilus rutilus were studied in relation to quality and quantity of food (Crans & Slaff 1977, Focks & Boston 1979). But there have been no studies on quantification of nutritional ecology of Toxorhynchites. Therefore, it is relevant to quantify parameters of nutritional ecology (= bioenergetics) of the Toxorhynchites that will help to overcome the problems associated with their mass-production and sustainable control. Nutritional ecology is the study of energy transformation in living systems. This information is essential for the proper interpretation of feeding, reproduction, defense, habitat selection and other life history phenomena in arthropods (Scriber & Slansky 1981). Lower larval survival has been reported at the highest feeding levels when Ae. aegypti was reared with excess of food in the laboratory (Arrivillaga & Barrera 2004). Starvation or nutritional deficiency is major cause of low egg production, deficiency of growth and sexual maturity. When food quality was changed, the rate of development, body composition and growth of arthropods differed significantly (Waldbauer 1968). If the conversion efficiency of insects were to differ depending on the type of food and its nutritional value, it would be of interest to know whether it is reflected in the growth pattern (Lawton 1971, Wigglesworth 1972, Cairns 1978, 1982). This study was carried out to find the effect of quality and amount of food offered on immature developmental duration, consumption rate and the relative growth rate of the larvae of T. splendens (Wiedemann). MATERIALS AND METHODS The experiment was conducted as a two-way design by offering four different instars of A. aegypti, A. stephensi (Liston) and C. quinquefasciatus (Say) to the predator at densities 10, 20, 40, 80 and 160 prey/predator/day. Amount of food offered in mg dry wt were treated as co-variate in the analysis. A wild type of laboratory strain of T. splendens bred from third and fourth instars collected from field in 1977 was used throughout the study and was in generation F182 at the time of experimentation. The required numbers of prey were placed in rectangular enamel trays, 45x40x5 cm (0.05 m2) containing 500 ml tap water with a single newly hatched larva of standard laboratory reared T. splendens. The number of prey consumed in 24 h period by a predator was estimated by subtracting the number alive from the total number offered. The remaining alive larvae were removed and fresh ones offered to the predator at respective densities. This was done at the interval of every 24 h until the predator larva pupated. Since climatic variation might influence feeding rate, all experiments were conducted in controlled temperature (25-27oC) and relative humidity (60-70%). Each test combination was replicated five times (n = 5). To estimate the amount of food consumed (mg dry wt) by the predator per day from the number eaten, dry weight of the prey was estimated. To determine the dry weight, thousand first and second instars and two hundred third and fourth instars of prey species were placed separately inside an oven with temperature maintained at 50oC for 24 h. The dried larvae were allowed to cool in desiccators and weighed with a Dhona chemical balance to the nearest 0.1 mg precision (Table I). Number of prey eaten by the predator was multiplied by their dry weight to get the amount of food consumed. Dry weight of the predator pupa was estimated following the same method. Estimated dry weight of the first instar predator was used for computing the weight gain (G). Weight gain is determined by subtracting the dry weight of the first instar from the dry weight of the pupa. In determining the total weight gain of the predator the error contributed by gut contents was negligible and may be neglected. Faeces collection was done by drying the beaker in desiccators and subtracting the previously weighted beaker from the one with dry faeces. Although temperature of the rearing medium is important influencing factor on the bioenergetics of the predator, its effect was not investigated but blocked out of the experiment by making the observations under identical temperature condition. From these measurements, following bioenergetics indices were calculated following Waldbauer (1968). The rate of food consumption (Cr) = Tc/(M xD) where Cr = amount of food consumed (mg dry wt/predator/day); Tc = total mass of food consumed (mg dry wt) during the entire immature development of a predator; M = mid body weight; D = duration required for completing the development. "Mid body weight" is defined as the weight gained from first instar to pupa divided by the duration of the development. Assimilation efficiency (Ase) (%) = [(C F)/C] x 100 where C = dry mass food consumed; F = dry mass of faeces egested. Relative growth rate (RGR) = In [(W2-W1)/(t2-t 1)] where W1 = dry weight of the first instar; W2 = dry weight of the pupa; t1 = age of the first instar (0); t2 = age of the pupa. RGR is the per day body weight gain of the predator. The effect of variables such as prey species, their instars and their interactions were studied by an analysis of co-variance in which food level was used as co-variate. Slopes of the regression were tested for homogeneity before performing ANCOVA on the data following the procedure of Sokal and Rohlf (1981). The coefficient of determination (r2) was computed for the linear regression of each set of data to determine the fit achieved by the regression. Immature duration and Cr were ln transformed before statistics being applied. Regression equations were tested for significant departure from zero using ANOVA (Sokal & Rolf 1981). Since immature duration and Cr of the predator differed in response to food level, significant differences among regression coefficients were tested by T'-method (Gabriel 1978) by setting 95% comparison intervals to slopes. The preliminary ANCOVA on RGR showed that groups were sampled from population of equal slopes. The results were displayed by setting 95% comparison intervals for means. The relationships between immature duration and Cr as well as Cr and RGR were studied by correlation analysis by using mean values of amount of food offered to four instars of three prey species. Similarly, the correlation between Cr and assimilation efficiency (Ase) was studied. The amount of protein, steroids, vitamins, and sugar in the food largely determine the developmental time, feeding rate and growth rate of an organism. Hence, to ascertain whether four instars of the three prey species differ significantly from each other in the protein content of their body, protein estimation was done following the method of Lowry et al. (1951). Protein estimation - One gm (wet weight) of second, third and fourth instars of the three prey species was homogenized separately in 10 ml of Tris-HCl buffer (pH 6.8) using a Potter's homogenizer. The homogenates were centrifuged at 5000 rpm for 10 min and the supernatants were stored at 4oC until use. The standard protein (BSA) solution was pippetted out into a series of test tubes in the range of 0.1, 0.2, 0.3, 0.4, and 0.5 ml to obtain 100, 200, 300, 400, and 500 µg respectively. The volume was made up to 2 ml with distilled water. A parallel tube with distilled water served as blank. The test solutions contained 0.1 ml extracts in 2 ml distilled water. To all the tubes, 4.5 ml alkaline copper reagent was added, mixed thoroughly and allowed to stand at room temperature for 10 min. Then, 0.5 ml diluted (2 times) Folin-Ciocalteau reagent was added rapidly with immediate mixing. After 30 min, the colour developed was read against the blank at 660 rm. The protein concentration of the test solutions was estimated after preparing the standard graph. The same procedure was followed for estimating protein of instars of prey species and each set was triplicated (n = 3). RESULTS The results of protein analysis showed significant difference in the amount of protein present (mg/gm body wt) among the prey species (Table II). The protein content of An. stephensi was significantly higher than Ae. aegypti and Cx. quinquefasciatus (F = 4.23; P < 0.05). However, there was no significant difference in the amount of protein present among the prey instars (F = 1.63; P > 0.05). The results of the qualitative and quantitative effects of food on bio-energetic parameters of the predator such as Id, Cr, assimilation efficiency (Ase), and relative growth rate (RGR) are given in Table III. The basic analysis of regressions separately for sets of data of Id, Cr, and RGR showed good fit in most of the cases (Tables IV, V, VI). Slopes were highly significant in all the cases of Cr (Figs 1, 2, 3). The slopes of RGR were significant in first, second and third instars of Ae. aegypti (Fig.4) and the first instars of An. stephensi and Cx. quinquefasciatus respectively (Fig.5). Results of regression of immature duration on food level indicated that immature developmental period of the predator decreased significantly with increase in food level (F = 2.96-34.69; P 0.001 < 0.05; Table IV). Since the slopes were unequal, the differences among slopes were tested following the method of Sokal and Rohlf (1981). The results showed that variation among nine regressions was highly significant (F = 7.35; df = 27; P < 0.001). There was an inverse relationship between the time taken by the predator to complete its immature development and the amount of food offered with type of prey (species and size) acting as influencing factors. Unplanned comparison among regression coefficients by setting 95% comparison intervals indicated that the degree of the slopes was not a function of prey species but differed among prey instars. The specific characteristic of Tx .splendens is that even without food (prey), it can withstand starvation and go for dormancy Results of regression of Cr on food level indicated that Cr of the predator increased significantly with increase in food level (Table V). Since the slopes were unequal, the differences among slopes were tested. The results showed that variation among twelve regressions was highly significant (F = 11.22; df = 36; P < 0.001). All the slopes of the regression were highly significant in the case of Cr with respect to four instars of three prey species offered as food (Figs 1, 2, 3, 4, 5). The results of regression of RGR on food level indicated that RGR increased significantly with increase in food level (Table VI). There were significant regressions of RGR on food level for first, second and third instars of Ae.aegypti as prey whereas the slope of the fourth instar was not significant (Fig. 4). Only the slopes of first instars of An. stephensi and Cx. quinquefasciatus were found significant (Fig. 5). Since the slopes were equal (F = 2.21; P > 0.05), differences among adjusted means were tested using the average regression co-efficient in a 2-way ANOVA (Sokal & Rohlf 1981). The common regression co-efficient was highly significant (F = 317.32; P < 0.001). Results of the significance tests indicated that there was a highly significant prey species and prey instar interaction (F = 13.36; P = < 0.001). Hence, differences in RGR depended on the prey species and prey instars. Multiple comparison by setting 95% comparison intervals to adjusted means showed significant increase in the RGR when An. stephensi was offered. The relative growth rate of the predator did not differ significantly when prey were either Ae. aegypti or Cx. quinquefasciatus. There was a significant decrease in RGR with first instar prey (F = 24.17; P < 0.001). However, RGR was equally more with second and third instar prey. The functional relationship between food level and Ase could not be established as there were no evidence of regression (P > 0.05). There was no significant correlation between Cr and Ase (P > 0.05). The Cr and RGR were correlated significantly in majority of the cases (P < 0.001 - < 0.01). Tests of homogeneity among correlation coefficients showed heterogeneity among the r's (χ2 = 66.52; df = 11; P < 0.001). So, Cr and RGR differed in their degree of relationship among prey species and prey instars. DISCUSSION The results suggest that both quality and quantity of the food influence the life characteristics of T. splendens during its immature development. However, the age of the predator could influence the food intake and subsequent growth rate. As the fourth instar of the predator is the gradual non-feeding stage, the regression models fit so poorly when compared to other instars of the predator. Increase in consumption rate resulted in reduction in immature duration. This is considered an adaptive character of the predator as this results in increase in population turnover in a reduced time when the prey is abundant. When prey was abundant there was an increase in the rate of predation. This resulted in reduced developmental time with increase in relative growth rate. The decreased growth rate of the predator with first instar prey might be due to high-energy utilization for capturing small prey. However, energy required for capturing the first instar of An. stephensi and Culex or Aedes may not be the same. This was more pronounced when the prey constituted larvae of An. stephensi as a result of more nutritional value of this prey species. Further, the encounter between the predator and the larvae of An. stephensi is ensured because of similarity in their mobility, use of foraging area and the position on the water surface (niche overlap) (James & Liston 1985). This could be a reason for high rate of predation on An. stephensi than on the other prey species. However, the co-occurrence of the larvae of these two species in the same containers is seldom observed in nature. The significant correlation between Cr and RGR indicates that the predator converts the food into body substance efficiently. The high protein content of the larvae of An. stephensi makes the processes of conversion of food into body substance energetically less expensive and therefore elevates RGR. Earlier studies showed that the quality of food consumed could influence the total body length and allometric growth of various organs through energetic-the conversion efficiency, conversion rate, assimilation efficiency and assimilation rate in the experimental population (Slanky & Scriber 1985). It was also suggested that the type of food had an impact on the life span and survival value of the animal (Madhavan & Bhaskaran 1975, Venkatesan & Raghunatha Rao 1979). The predator compensated the loss in daily food intake at low food level by extending its immature duration thereby increasing its total food intake during its development period. This is an important adaptive character for the predator to cross the minimum threshold pupal weight for adult emergence during scarce prey availability. Studies of Lounibos et al. (1979) showed that these adaptations of a predator would help in increasing its population turnover in a reduced time when the prey is abundant. Further, they added that the prolongation of developmental time of aquatic stages of insect predators at the time of less prey availability would help the predator to acquire the minimum energy reserve for attaining the minimum threshold pupal weight required for adult emergence. This physiological adaptation helps the predator population to survive till the time of availability of abundant food. Growth in animals involves conversion of extraneous matter into live tissue through the process of ingestion, absorption, assimilation and synthesis. Besides energy spent in capturing the prey, conversion of food into body substance requires energy. The balance of energy that remains after these expenses is used in growth. The loss in daily food intake at low food level is therefore compensated by delay in the pupation. This is found to be an another important adaptive character of the predator to acquire the minimum energy reserve for the full development of gonads in the adults (Marshall & Kevan 1962, Murdoch 1966, Campbell & Sinha 1978). The results show that T. splendens is well adapted to continuous insectary conditions. It has evolved different life characteristic strategies such as difference in the time taken to become pupa, different feeding and growth rates in response to food availability both in quality and quantity. These biological characteristics help in mass production of the predator for field releases. Developmental period depends on number and nutrient value of prey offered to the predator. Such an active feeding on more protein diet helps in storing more nutrients and energy to tide-over the non-feeding pupal and comparatively less active feeding adult period. Nutrient reserve also increases fecundity and survival of the adult (Dominic Amalraj & Das 1996). This facilitates the production of larva number of T. splendens for field release at the time of onset of monsoon prior to increase in Ae. aegypti population. Conversely, during other periods of the year, T. splendens could be maintained in less number on less prey diet. Since T. splendens is found to prolong its immature duration at times when prey is less in number, the generation time is extended till the time of availability of enough food. The overall significant negative correlation between immature duration and Cr (r = - 0.89, df = 60, p < 0.001) was expected since increase in daily food intake will accelerate the instar development thereby reducing the duration of the immature development, resulting in reduced generation time. When abundant prey becomes available, the generation time is reduced. This facilitates mass-rearing of the predator in large numbers. The ability of the predator to survive at low feeding rate (Crmaint) is an appreciable adaptation to tide over the period of the low prey availability. ACKNOWLEDGEMENTS To MP Prasad and K Sathianathan for their technical assistance. To Ms R Sundarambal for the literature survey. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05180t4.jpg] [oc05180f2.jpg] [oc05180f1.jpg] [oc05180t3.jpg] [oc05180t2.jpg] [oc05180t6.jpg] [oc05180t5.jpg] [oc05180f4.jpg] [oc05180f5.jpg] [oc05180t1.jpg] [oc05180f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}