 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
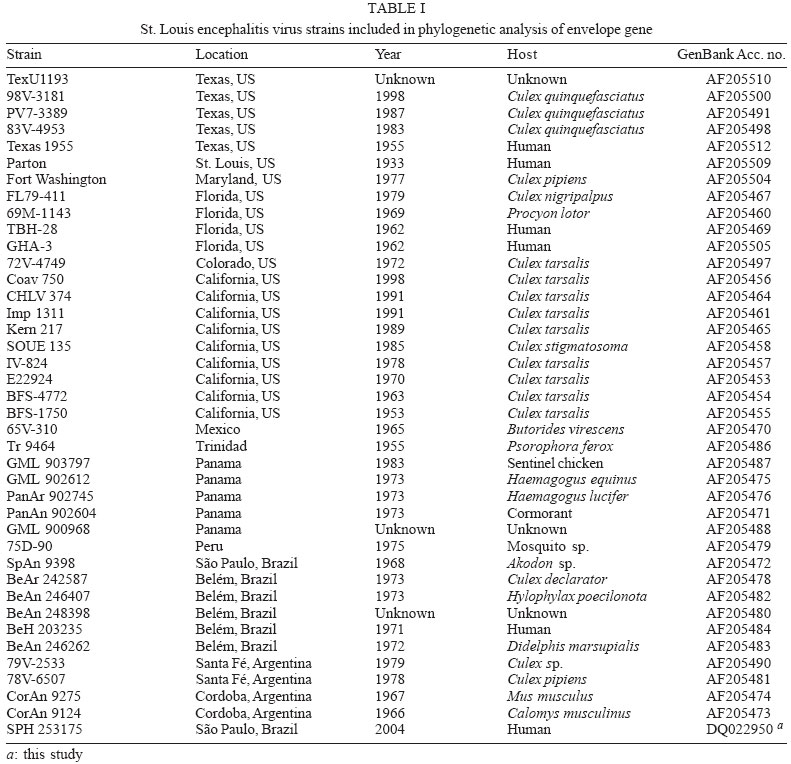
Memórias do Instituto Oswaldo Cruz, Vol. 101, No.1, February 2006, pp. 57-63 Genetic characterization of St. Louis encephalitis virus isolated from human in São Paulo, Brazil Cecília Luiza Simões dos Santos/+, Maria Anice Mureb Sallum*, Heitor Moreira Franco, Fabíola Maiumi Oshiro, Iray Maria Rocco Serviço de Virologia, Instituto Adolfo Lutz, Av. Dr. Arnaldo 355, 01246-902 São Paulo, SP, Brasil *Departamento de Epidemiologia, Faculdade de Saúde Pública, Universidade de São Paulo, São Paulo, SP, Brasil Financial support: Fapesp (no. 2004/07819-6) and CNPq (no. 305339/2003-6); to CLSS; to MAMS CNPq (501529/2003-9) +Corresponding author: simoes.santos@uol.com.br Received 29 August
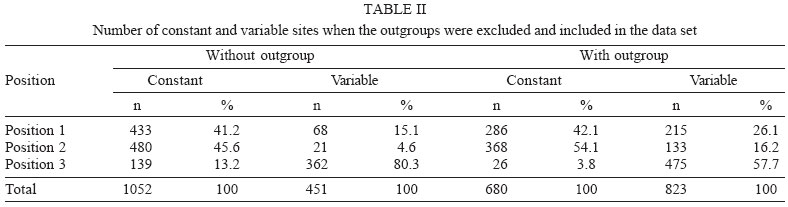
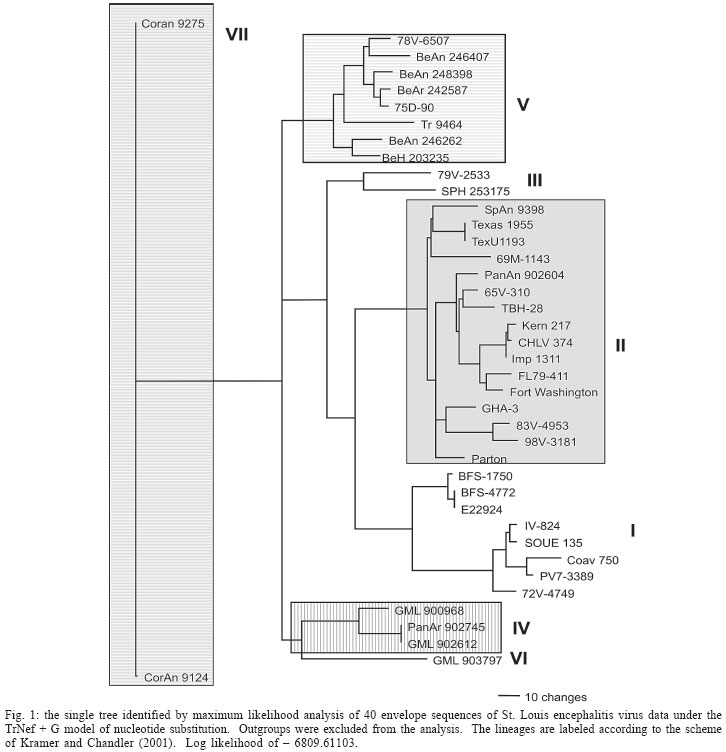
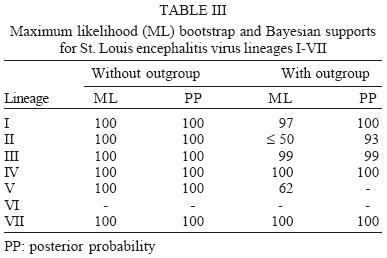
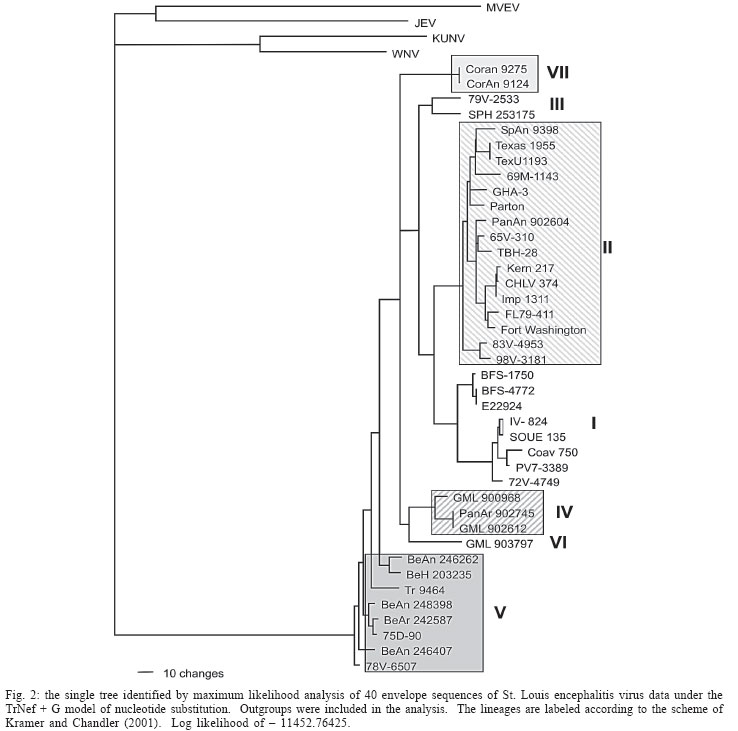
2005 Code Number: oc06011 The molecular characterization of SPH253157, a new strain of St. Louis encephalitis virus (SLEV), isolated in 2004 from the first case of human infection recognized in the state of São Paulo, Brazil, is reported. The patient, presenting a febrile illness without neurological involvement, was hospitalized as a probable case of dengue fever. Genomic RNA was isolated from the supernatant of C6/36 cells infected with acute phase-serum specimen of the patient and the envelope gene was amplified by reverse-transcription-polymerase chain reaction. The complete nucleotide sequence of the envelope gene of this isolate was directly sequenced from the amplified products and compared with other Brazilian and American SLEV strains. Phylogenetic analyses were carried out under maximum likelihood criterion with outgroups both included and excluded. Outgroups comprised four flavivirus of the Japanese encephalitis group. Phylogeny also included Bayesian analysis. The results indicated that the new SLEV isolate belongs to lineage III, being closely related to an Argentinean strain recovered from Culex sp. in 1979. It is concluded that there are at least 3 lineages of SLEV in Brazil. Key words: St. Louis encephalitis virus - envelope gene sequence - phylogeny - Brazil St Louis encephalitis virus (SLEV) belongs to the family Flaviviridae, genus Flavivirus, within the Japanese encephalitis virus group (Calisher et al. 1989). The genome of flaviviruses contains a single-stranded positive sense RNA, about 10,800 nucleotides long that encodes 10 distinct proteins in a single reading frame (Chambers et al. 1990). SLEV is the etiologic agent of St. Louis encephalitis, a disease of epidemiological importance in North America. This virus was first isolated in the United States of America (US) in 1933, during an outbreak, which occurred in St. Louis and Kansas City, Mo. (Muckenfuss et al. 1934). In the US intermittent and unpredictable epidemics have been occurring during the last decades. The majority of SLEV infections is sub clinical or result in mild illness. The severity of infections is strongly dependent on the age of a patient, and the case fatality rate during epidemics ranges from 2% in young adults to more than 22% in the elders (Monath & Heinz 1996). In spite of being endemic in Central and South America, causing sporadic illness, occurrence of SLEV outbreaks is unknown (Spence 1980, Monath & Heinz 1996). The biological cycle of SLEV in nature involves primarily wild birds and several mosquito species; humans and other mammals are considered to be incidental hosts (Monath & Heinz 1996). In the US, Culicinae mosquitoes and birds, Passeriformes and Columbiformes, are involved in the dynamics of SLEV transmission. The transmission cycle varies regionally, depending on the biology of mosquito species, virulence of SLEV strains and susceptibility to infection of vertebrate hosts (Monath & Tsai 1987, Day 2001, Reisen 2003). In Central and South America SLEV transmission cycle is not well defined, but it may include mosquito species of the genera Culex, Mansonia, Haemagogus, and Sabethes. A variety of bird taxa, including herons, egrets, and cormorant may be virus reservoirs (Spence 1980, Monath & Heinz 1996, Reisen 2003). The occurrence of SLEV in Brazil was recorded in 1953, by the finding of antibodies to this virus in residents of the Amazon Valley (Causey & Theiller 1958). A pool of mosquito Sabethes belisarioi, captured along the Belém-Brasília highway, was the source for the first isolation of a SLEV strain (Causey et al. 1964). Later in 1970 and 1978 two strains were recovered from human blood in Amazon region. In both cases the patients developed a febrile illness with jaundice, neither of them presenting neurological involvement (Pinheiro et al. 1981, Vasconcelos et al. 1998). SLEV was also found in the Southeast region of Brazil where it was isolated from wild birds, rodents, and sentinel mice during a surveillance program on arbovirus activity carried out in the state of São Paulo from 1967 to 1969 (Lopes et al. 1979). In the summer of 2004, after more than two decades of the last human Brazilian report, a clinical case of infection by SLEV was detected in the state of São Paulo. Epidemiologic data about the SLEV case was fully detailed by Rocco et al. (2005). Briefly, a patient, suffering from severe uncharacterized febrile illness without neurological involvement, was hospitalized as a probable case of dengue fever. Serological tests detected antibodies to fla-vivirus but were negative in tests specific for Dengue virus (DENV). The virus was isolated in cells culture inoculated with patient acute phase serum and identified by reverse-transcription-polymerase chain reaction (RT-PCR) and sequencing (Rocco et al. 2005). SPH 253157 is the first strain of SLEV isolated from human reported in the state of São Paulo and the third one in Brazil. In Brazil, the genetic diversity of SLEV has been poorly explored and only a few strains have been characterized at molecular level. In the present study, we determined the entire nucleotide sequences of the Envelope (E) gene of SPH 253157 strain. In addition, representative E sequences of SLEV lineages as well as previously published E sequences of Brazilian SLEV were downloaded from GenBank (Kramer & Chandler 2001) and included in the study for comparison with SPH 253157 strain. The main objectives of this study are: (1) to estimate genetic relationships among SPH 253157 and several previously collected Brazilian strains and other SLEV American isolates; (2) to characterize genetic lineage of SPH 253157 isolated from a human patient infected in the state of São Paulo. MATERIALS AND METHODS Viruses - The SLEV SPH 253157 strain was isolated from acute phase-serum specimen of a patient in monolayer cultures of clone C6/36 cells of Aedes albopictus (Rocco et al. 2005). Viral strains, locations, years of isolation, and GenBank accession numbers of the E sequences utilized in the phylogenetic analyses are listed in Table I. Sequences from Japanese encephalitis virus (JEV) (Accession # M55506 (Nitayaphan et al. 1990); West Nile virus (WNV) Accession # M12294 (Castle et al. 1985); Kunjin virus (KUNV) Accession # D000246 (Coia et al. 1988), and Murray Valley encephalitis virus (MVEV), Accession # X03467 (Dalgarno et al. 1986) were used as outgroups in the phylogenetic analysis. RT-PCR and nucleotide sequencing - Viral RNA was isolated from first passage cell culture supernatant using the QIAmp Viral RNA Extraction Kit (QIAGEN, Valencia, CA, US), according to the manufacturer's instructions. Primers designed to amplify the complete E gene sequence, described by Kramer and Chandler (2001), were used in RT-PCR and sequencing reactions. RT-PCR was performed with the SuperScriptTM One-Step RT-PCR with PlatiniumR Taq System (Invitrogen/Life Technologies, CA, US) in the presence of 4 µl of viral RNA and 10 picoMol of each primer. RT-PCR products were directly sequenced using the "ABI PrismR Big DyeM Terminator Cycle Sequencing Ready Reaction Kit" (PE Applied Biosystems, Foster City, CA, US), according to the manufacturer's protocol. Sequences were determined in 377 ABI sequencer (PE Applied Biosystems). The E gene sequence of SPH 253157 is deposited in GenBank Data Libraries under accession number DQ022950. Phylogenetic analysis - Sequence alignment was performed using the multiple sequence alignment method implemented in CLUSTALX (Thompson et al. 1997). Accuracy of nucleotide sequence alignment was examined using amino acid sequence alignment. Maximum likelihood (ML) analyses were performed using PAUP 4.0b10 (Swofford 2003). The best model in PAUP was chosen using ModelTest 3.06 (Posada & Crandall 1998). This program uses both a hierarchical Likelihood Ratio Test (hLRT) and the AIC (Akaike Information Criterion) to choose among available models; when the hLRT and AIC disagreed the simpler model was chosen. Maximum likelihood searches started with a neighbor-joining tree, on which we then optimized parameters and fixed the values for those parameters for branch swapping on that tree. For each bootstrapped data set PAUP did TBR branch-swapping, and only one tree was saved in each replicate (multrees = no). Support for each clade generated for unpartitioned data sets was assessed by 1000 bootstrap replicates using PAUP. For Bayesian analysis, we used the program MrBayes (Huelsenbeck & Ronquist 2001). Since the kinds of models available in this program have limited overlap with those in PAUP*, Bayesian analysis was carried out using GTR plus site-specific model. Program default values for prior probabilities were used. The Markov Chain Monte Carlo (MCMC) was 1,000,000 generations long, sampling every 100 generations, for a total of 10,001 samples. Of these, the first 1000 were discarded as burn-in, which is well past the point where the likelihood plot reached a plateau. Evidence for recombination events - Recombination exploratory tests were performed using the SimPlot program, version 2.5 (Lole et al. 1999). Bootscanning analysis (Salminen et al. 1995) was used to evaluate genetic relatedness between the query sequence and a panel consisting of 39 sequences from SLEV lineages used in the study. A sliding window of 400 bp was moved across the alignment in 20 bp steps, with gap strip on. The genetic distance among sequences in each window, based on neighbor-joining method under Kimura 2-Parameters model was estimated using 100 bootstrap replicates. RESULTS Nucleotide and amino acid sequences analyses - Comparison of the nucleotide and amino acid E sequences determined for the strain SPH 253157 with those from representative SLEV American isolates showed a high level of identity. The paired identity ranged from 89.4 to 100% (nucleotide) and 97.4 to 100% (amino acid) (MEGALIGN, DNASTAR, Inc.). The strain with the highest percent of nucleotide similarity in relation to SPH 253157 was 79V-2533, an Argentinean isolate recovered in 1979 from Culex sp. mosquitoes. Phylogenetic analysis - Considering both the ingroup and outgroups, the alignment of the E gene sequences generated 1503 sites. The number of constant and variable sites when the outgroups were both included and excluded are in Table II. Alignments are available on request. A primary objective of this study was to characterize the lineage of the strain SPH 253175 and consequently to test how many phylogenetic lineages exist whitin the group (SLEV). Since the outgroup was quite divergent we suspected that it might cause us to choose inappropriate models or be responsible for long branch effects. To test this, all the analyses were conducted with and without the outgroups. Phylogenetic analysis with the outgroups excluded - ML (Fig. 1) and Bayesian (not shown) topologies were generally the same. They recovered all 40 sequences included in data set into seven phylogenetic lineages, generally according to their geographic origin. These lineages are similar to those identified by Kramer and Chandler (2001). ML analyses were carried under the TrNef + G model suggested by ModelTest. The ML topology is shown in Fig. 1. SPH 253175 sequence clustered together with 79V-2533 strain of lineage III of Kramer and Chandler (2001). Bootstrap support for the split leading to lineage III is 99%. This lineage is clustered in a major, weakly supported group consisting of lineages I and II (Table III). ML bootstrap support for separate groups was generally moderate to strong, however the split leading to lineages IV and VI is weakly supported (≥ 50%) and the branch separating the entire lineage I from lineage II is moderately supported (85%) (Table III). Bayesian 50% majority rule consensus tree (not shown) is similar to ML topology generated under TrNef + G model. Posterior probabilities for relationships among SLEV lineages are generally higher than ML bootstrap values (Table III). Phylogenetic analysis with the outgroups included - For the ML analysis, ModelTest suggested the TrNef + G model, the same model suggested when the outgroups were excluded. The position of the root has 99% bootstrap support. However, maximum likelihood support for the splits leading to lineages II and V is weak (Fig. 2). For the Bayesian analysis, a site-specific model was used. The settings were as described above for the data without the outgroups. The support for relationships among the sequences of each ingroup lineage was generally lower when the outgroups were included in both ML and Bayesian analyses than when the outgroups were excluded (Table III). Evidence for recombination events - A search for recombination events, in the strain SPH 253157 was performed by bootscanning analysis (Salminen et al. 1995) over the E sequence. The results failed to reveal any recombinant segments in that genome region (results not shown). DISCUSSION All our results revealed that the strain SPH 253157 recovered from a human patient in the 2004 summer is distinct from all six previously characterized Brazilian SLEV. It clusters in lineage III of Kramer and Chandler (2001), which includes Argentinean 79V-2533 strain. Other Brazilian strains isolated from distinct hosts from 1968 until 1973 grouped with other South American and Trinidad samples in lineage V. One Brazilian strain isolated from Akodon sp. clustered with several US isolates in lineage II. Consequently, three distinct lineages of SLEV have been reported in Brazil, suggesting that the virus is circulating and dispersing throughout the Americas. This hypothesis is supported by a recent finding of Twiddy and Holmes (2003) who demonstrated strain GMO94, isolated in Guatemala, to be a recombinant of South (CorAn 9124, Cordoba, Argentina) and North (TNM-711K, Tennessee, US) American strains. Results of the present phylogenetic analyses corroborate Kramer and Chandler's (2001) hypothesis that the lineages of SLEV virus, which are circulating in the Americas, belong to distinct lineages, and that exists a geographic specificity. Consequently the dynamics of SLEV transmission may be distinct, with the involvement of distinct vectors and virus reservoirs. Results of studies carried out in the Brazilian Amazon region found Culex declarator and Culex coronator to be infected with SLEV. Additionally, members of the families Formicariidae (Formicarius analis, Conopophaga aurita, Tham-nomanes caesius and Hylophylax poecilonota), Pipiridae (Chiroxiphia pareola) and Columbidae (Geotrygon montana) were sources of viral isolates and could be involved in the maintenance of the SLEV cycle in that region. Furthermore, monkeys, armadillos, and marsupials may be vertebrate amplifying hosts (Vasconcelos et al. 1998). Regarding to the dynamics of the transmission of SPH 253157 strain isolated in São Paulo, it remains unclear because no virus was isolated either from birds or mosquitoes during the eco-epidemiological studies carried out in that region where the patient was probably infected (Rocco et al. 2005). It is important to emphasize that infection caused by SLE virus, as well as several arboviruses that affect the central nervous system, presents diverse symptoms that are hardly diagnosed unless in cases of epidemics. Few physicians suspect initially of the disease even in endemic areas (Reisen 2003). In the present human case of SLEV infection, it was first suspected as a dengue case. The following epidemiological investigation included clinical and laboratorial studies which led to the successfully identification of SLEV as the etiological agent. The lack of systematic epidemiological surveillance activities toward arboviruses of minor importance compared to Yellow fever, Dengue fever, and Rocio encephalitis, for instance, may partially explain the few number of clinical cases of Saint Louis encephalitis reported both in Brazil and tropical America. Others factors including the attenuation of viral strains that was observed in rodent strains isolated in São Paulo and Argentina (Monath et al. 1980) and the early acquisition and accumulation of human immunity, revealed among residents in the Amazon region (Pinheiro et al. 1986, Vasconcelos et al. 1998) and in different areas of the state of São Paulo (Iversson et al. 1981, Romano-Lieber & Iversson 2000) as well as in several Argentina counties (Sabattini et al. 1998) may be also considered. Finally, a particular concern is relative to WNV, another member of the Japanese encephalitis group, which transmission cycle involves wild birds and Culex mosquitoes. WNV caused an outbreak of encephalitis in New York in 1999 and SLEV was initially suspect to be the etiological agent (Briese et al. 1999, Campbell et al. 2002). Lillibridge et al. (2004) reported that in Harris County, Texas, an endemic region for SLEV, WNV appeared and spread rapidly throughout the area. Surveillance data indicated that SLEV and WNV can coexist, despite their ecologic, antigenic and genetic similarities and both viruses will probably persist in Harris County. Consequently, it is important to emphasize the necessity of robust surveillance program for arboviruses, which are generally weak or even inexistent in several countries (Gubler 2002). Furthermore, to reduce the risk of epidemics, health professionals, especially in cases of hardly recognized viral infections, should carry out exhaustive clinical, epidemiological and laboratorial investigations. Diagnostics methods as those represented by reverse-transcription amplification techniques, including the recent real time-PCR procedure (Mackay et al. 2002) are specially recommended to ensure rapid identification of the viral agent and allow a prompt surveillance action by health authorities. ACKNOWLEDGEMENTS To Dr Tiyo Sakurai (Instituto Adolfo Lutz, São Paulo, Brazil) and Dr Peter G Foster (Natural History Museum, London, UK) for suggestions and providing helpful comments to improve the paper. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06011f1.jpg] [oc06011t2.jpg] [oc06011t1.jpg] [oc06011f2.jpg] [oc06011t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}