 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 101, No. 3, May 2006, pp. 269-272 Localization of Brugia malayi (sub-periodic) adults in different organs of Mastomys coucha and its influence on microfilaraemia and host antibody response K Athisaya Mary+, SL Hoti, KP Paily Vector Control Research Centre, Indian Council of Medical Research, Indira Nagar, Pondicherry-605 006, India +Corresponding author: athisaya@yahoo.com Received
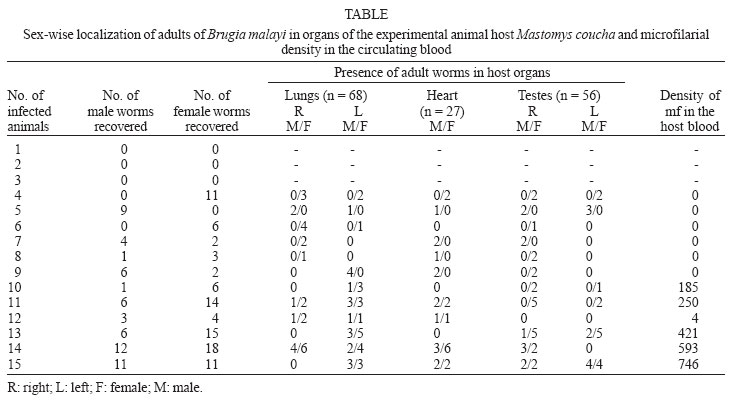
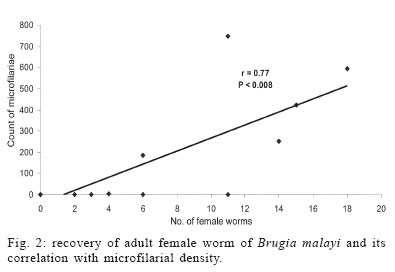
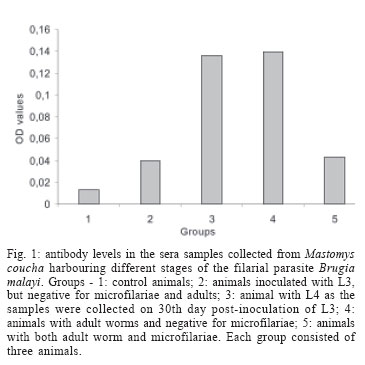
4 November 2005 Code Number: oc06047 Lymphatic filariasis caused by nematode parasites Wuchereria bancrofti or Brugia malayi is a spectral disease and produces wide range of immune responses and varying levels of microfilaraemia in infected individuals. The relationship between the immune response of host and the developmental stage of the parasite as well as the microfilariae (mf) density and specific location of the adult worms is yet to be understood. As an experimental model, B. malayi adapted in the experimental animal Mastomys coucha has been used widely for various studies in filariasis. The present study was to assess microfilaraemia as well as the humoral immune response of M. coucha during various stages of B. malayi development and their localization in different organs. The result showed that the density of mf in the circulating blood of the experimental animal depended upon the number of female worms as well as the location and co-existence of male and female worms. The mf density in the blood increased with the increase in the number of females. The clearance of inoculated infective stage (L3) or single sex infection or segregation of male and female to different organs of infected host resulted in amicrofilaraemic condition. With respect to antibody response, those animals cleared L3 after inoculation and those with adult worm as well as mf showed low antibody levels. But those with developmental fourth stage and/or adult worms without mf showed significantly higher antibody levels. Key words: Mastomys coucha - Brugia malayi - adult worm - clearance - microfilariae - antibody levels Lymphatic filariasis is one of the major tropical diseases and is caused by filarial nematodes such as Wuchereria bancrofti, Brugia malayi, and Brugia timori. About 120 million people are either infected or with lymphatic pathology worldwide and among them India accounts for 42.8% of the global burden of the disease (WHO 2002). Information on the parasite build-up and host immune responses against different life stages of the filarial parasite are important to find out the ways and means of diagnosing the filarial infection and for other related studies. Mastomys coucha is the known laboratory animal model of B. malayi sub-periodic strain and 71-81% of this animal could develop the inoculated infective stage (L3) to the adult worms (Petranyi et al. 1975, Murthy et al. 1983). Despite the development of the worms, the status of host microfilaraemia, i.e., the presence and density of microfilariae (mf) in the circulating blood, is determined by various factors. For example, segregation of male and female due to their localization in different organs, or all the worms developing to single sex lead to non-production of mf (Paily et al. 1995). Also, these animals can exhibit a variety of immunological response depending upon the presence or absence of mf, their developmental stages, adults or their products (Rao & Klei 1997, Athisaya Mary et al. 2005). There are reports of human hosts infected with filarial parasites but not progressing to micro-filaraemic stage (Dreyer et al. 1996) and the reason for this could be the same as stated above. In the present study, M. coucha inoculated with L3 of B. malayi sub-periodic strain were examined for mf, organ-wise adult worm localization, and immunological status to find out whether there is any relationship between these factors. MATERIALS AND METHODS Infection of animal and monitoring microfilaraemia - Adult worm localization, microfilaraemia, and host immune response was studied using male M. coucha inoculated with L3 stage of B. malayi sub-periodic strain (Ash & Riley 1970). Five groups of 3 male animals each were inoculated subcutaneously (s/c) with L3 harvested from 1200 infected Aedes aegypti (Liverpool strain), by standard procedure (Lim & Sim 1983). At the time of inoculation, the animals were 6-8 weeks old and a single animal received 100 L3 suspended in 0.5 ml of saline (0.9%). Animals were inoculated serially and monitored periodically, at weekly intervals, from day 90 post-inoculation of L3 until blood presence of mf and their density in the circulating blood. This was done by drawing 20 mm3 of tail vein blood onto a micro slide and making into blood smear. The blood smears (5/animal) were dehaemoglobinized, stained with Giemsa's stain, and examined under a compound microscope at a magnification of 100 x for the presence and count of mf. The relationship between mf density and number of female worms was checked by least square regression analysis. Blood samples of 0.5 to 1 ml/animal was collected from the tail vein on 30th day (corresponding to the presence of L4 stage larvae of the parasite in the host animal) and 6 months (just before necropsy of the animal), sera was separated and stored at 20ºC until further use. Recovery of adult worms from infected animal - The animals were necropsied after 6 months to recover the adult worms localized in different organs (Murthy et al. 1995). For this, the organs such as lungs, heart, and testes dissected out from animals were placed in separate cavity blocks, teased out in normal saline (0.85%) and kept for 1 h for the worms to come out. The number of worms recovered from each organ was recorded sex-wise after examining them under a compound microscope at a magnification of 40x. Assessment of antibody levels in animals with different stages of the filarial parasite - Antibody levels in the sera samples collected from the experimental animals (n = 15) on 30th day and 6th month was determined in replicates by direct-ELISA. Different stages of the B. malayi such as the mf, L3, L4, and adult were obtained from a colony of the parasite maintained in jird, Meriones unguiculatus, as the mammalian host and A. aegypti as the mosquito host. Antigens of these stages were prepared as follows: each of the parasite stage (2 x 105 mf, 1800 L3, 65 L4, and 20 adult female) was homogenized in phosphate buffered saline (PBS, pH 7.4) in a tissue homogenizer (Top syringes Ltd., Mumbai, India) and the homogenate was sonicated (Ultrasonic processor XL, Heat systems, Denmark) at 13 Hz for 8 cycles of 30 s each, in the presence of a mixture of protease inhibitors such as EDTA-1 M, EGTA-1 M, NEM-1 M, Pepstatin-1 mM, PMSF-0.33 M, and TPCK-0.1 M at 5 µl/ml. After sonication, the homogenate was held overnight at 4ºC and centrifuged at 5000 rpm for 5 min in a refrigerated centrifuge. Protein concentration of the supernatant was estimated by the method of Lowry et al. (1951) and stored at 20ºC until further use. The antibody levels in the sera samples from the experimental animals were determined by direct-ELISA following standard procedure (Voller et al. 1976). Briefly, the antigens prepared from different stages of B. malayi were coated onto 96 well polysterene plates (Nunc, US) to have 0.5 µg of protein/well and incubated overnight at 4ºC. Sera samples from the experimental animals (diluted 1:100) were reacted for 2 h after blocking the wells with 1% bovine serum albumin (BSA). Anti-mouse peroxidase conjugate (Sigma Chem. Co., US) was allowed to react with the antigen-antibody complex, and q-Phenylenediamine dihydro-chloride (Sigma) was added as a chromogen in a substrate solution after another wash. After 20 min of incubation the reaction was arrested with 5 N H2SO4 and the optical density (OD) was read at 492 nm in an ELISA reader (Titertek Multiscan., Lab systems). Test sera samples with OD values greater than 2 Standard Deviations above the mean OD values obtained from the replicates of same dilution of sera samples of control animals (n = 3) inoculated with only PBS were considered as positive for antibodies (Saroj & Renapukar 1995). One way ANOVA was carried out to compare the difference in antibody levels of the sera samples of the different groups of animals. Pair wise comparison of antibody levels was done using the post-hoc multiple comparison test based on LSD. P value < 0.05 was considered as significant. RESULTS AND DISCUSSION The present study was carried out to find out the relationship between localization of adult filarial worms in different organs of the experimental animal, their effect on microfilaraemia and the stage wise immune response of the host in terms of the level of total IgG. The animal model M. coucha was used for infection and development of B. malayi, as considerable amount of information is available on the parasite life cycle in this model (Petranyi et al. 1975, Murthy et al. 1983, 1992, Tyagi et al. 1998). In this animal model, the pre-patent period was found to be around 111 days and the microfilaraemia persisted beyond 6 months. About 71 to 81% of the L3 inoculated male animals could develop the parasite to become microfilaraemic and 11% of the L3 developed to adult worms. The numbers of adult worms recovered from different organs of the experimental animals are presented in the Table. Out of 151 worms recovered from experimental animals (n = 15), 59 were males and 92 were females. Worms were distributed in organs like lungs, heart, and testes. More number of worms were recovered from lungs (n = 68) followed by testes (n = 56) and then the heart (n = 27). The number of female worms recovered from an animal ranged from 2-18, whereas that of male ranged from 1-12. A high rate (80%) of the animals showed adult worms and 50% of them were positive for mf. The mf density ranged from 4 to 746/20 mm3 of blood. Recovery of adult worms showed that the percentage of L3 developed to adult was low and it ranged from 4 to 30%. Three out of 15 animals inoculated with L3 stage failed to develop them to adult worms, even after 6 months as they were negative for both mf and adult worms. Similarly, there were animals (n = 3) with only female or male worms which remained mf negative throughout the study period and they could be termed as single sex infection. Few animals (n = 3) which harboured both sexes of the worms also did not show mf in the circulating blood as the male and females were segregated in different organs. This is because the inoculated L3 might have migrated to different organs of the animal to get localized and thus, remaining in isolated condition without mating. This is in accordance with the study conducted by Vincent et al. (1976), where the development of B. malayi in jird model M. unguiculatus showed a characteristic tendency to localise within the heart and pulmonary arteries. Animals with at least one male and one or more female adult worms residing together in one location showed varying density of mf depending on the number of females. The mf density increased significantly with increase in the number of female worms. There was a positive correlation (r = 0.77, df = 527, P < 0.008) between the number of female worms and the mf density (Fig. 2). The results thus showed that the clearance of inoculated L3 or, single sex infection or no meeting of male and female in an infected host could result in amicrofilaraemic state of infected animals. Unlike in animals, where the worms got lodged in major organs such as lungs, heart, and testes, the situation in human lymphatic filariasis is different as most of the worms develop in the lymphatics (Mak 1983). Still there are chances of segregation of male and female worms at different levels, in various lymphatics, and it could lead to different disease pattern and microfilaraemia. After getting the data on mf density and adult worm recovery from individual animals, they were grouped into (i) L3 inoculated, mf and adult negative, (ii) adult positive and mf negative, (iii) adult positive and mf positive. The means of serum antibody levels of animals belonging to these three groups are presented in Fig. 1. Animals inoculated with L3 stage, but failed to develop them showed low level of antibody and the mean OD value was 0.042 (SD = 0.004). It indicates that the L3 inoculated as well as the antibodies produced against it would have got cleared in due course of time or the L3 specific antibodies produced in the host's system would not have reacted with the adult stage or its excretory/secretory (e/s) antigens because of stage specificity of these antibodies. Similarly, animals with both adult worm as well as mf showed low antibody levels (mean OD value 0.040, SD = 0.001). It could be due to the downregulation of immune system as reported in human microfilaraemic individuals (King & Nutman 1991) where the release of successive broods of mf into the blood lead to the suppression of antigen molecules released and made them non-reactive (Murthy et al. 1995). However, sera samples collected on 30th day from animals that harboured L4 stage parasite or on 120th day, when the animals harboured adult stage parasite, but not mf, showed high antibody levels. The antibody levels in mean OD value was 0.137 (SD = 0.002) and 0.138 (SD = 0.004), respectively and it was significantly higher (F = 1134.942, df = 420, P < 0.000) than those of the earlier two groups. This may be due to the higher titre of antigens present in the animals with developing or adult parasites (Cheirmaraj et al. 1991) or could be due to the specificity of the antibody to antigen used for the assay. But the number of adult worms recovered and their location did not show any effect on the level of antibodies. Hence, the stage of the parasite rather than the number of worms plays an important role in deciding the level of antibodies in the infected host. Investigations on these antigens, which induced higher level of antibodies, might lead to better understanding of the stage specific immune response of the host during various stages of development of the filarial parasite. ACKNOWLEGEMENT To Dr PK Das and Dr K Balaraman, Vector Control Research Centre, for their encouragement and constant support. To Dr S Subramaniam, for the statistical support, to Mr A Ramamoorthy, for technical support. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz |

{kind=link}
{kind=link}
{kind=link}