 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 101, No. 3, May 2006, pp. 281-286 Antimicrobial susceptibility, phage types, and pulsetypes of Salmonella Typhimurium, in São Paulo, Brazil Ângela Cristina Rodrigues Ghilardi+, Ana Terezinha Tavechio, Sueli Aparecida Fernandes Seção de Bacteriologia, Instituto Adolfo Lutz, Av. Dr. Arnaldo 351, 9o andar, 01246-902 São Paulo, SP, Brasil +Corresponding author: aguilard@ial.sp.gov.br Received
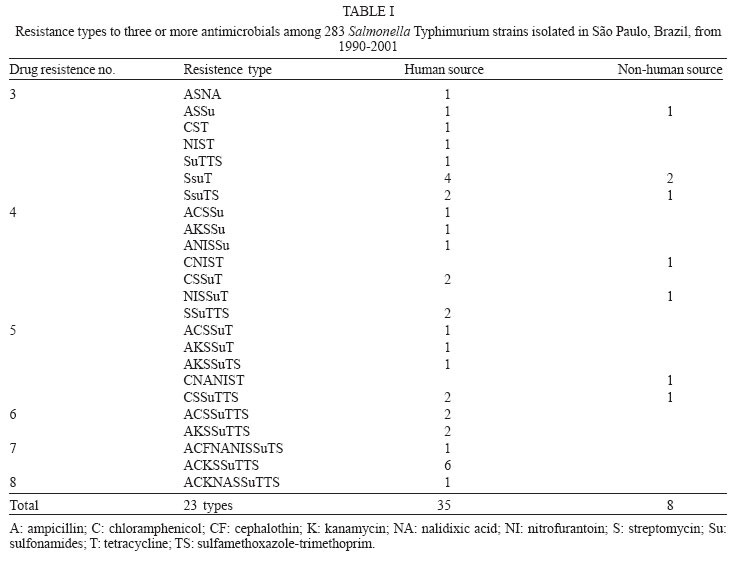
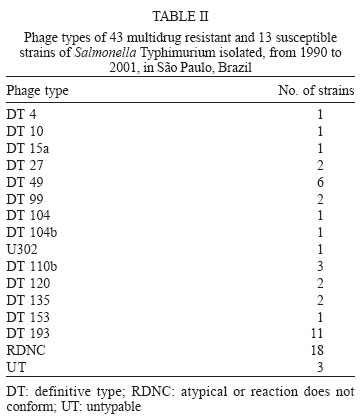
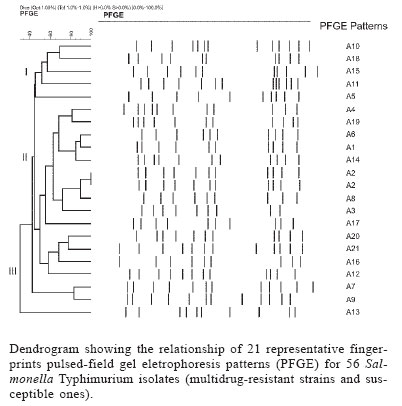
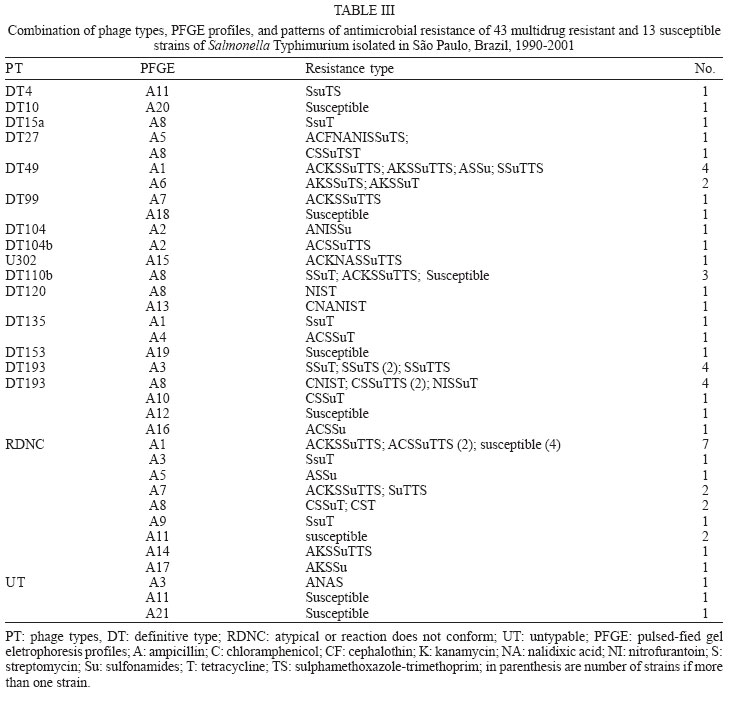
6 December 2005 Code Number: oc06043 A total of 283 Salmonella Typhimurium strains isolated from cases of human infections and non human sources, were examined for antimicrobial susceptibility and the incidence of resistance was 38% and multiple resistance (to three or more antimicrobials) was 15%. All 43 multidrug-resistant strains (MDR) and 13 susceptible ones were characterized by phage typing and pulsed- field gel electrophoresis (PFGE). The strains encompassed 14 definitive phage types (DT), three were untypable (UT), and 18 atypicals or reaction does not conform (RDNC), which belonged to 21 PFGE patterns, A1-A21. The predominant phage types were DT49, DT193, and RDNC and two strains belonging to DT 104 and 104b were also identified. The most commum PFGE patterns were A1 and A8. Analysis by PFGE and phage typing demonstrated that the most of the MDR were multiclonal and association among multiresistance, phage typing, and PFGE patterns was not so significant. Key words: Salmonella Typhimurium - phage types - multidrug-resistant Salmonella is one of the most important causal agents of foodborne illness. It can be isolated from a variety of foodstuff such as raw meats, poultry and poultry products, milk and milk products (Gorman & Adley 2004). The zoonotic nature of Salmonella infections is well established and the observed increase in human cases has been associated with high levels of infections in animal husbandry, mainly in pigs and poultry (Seyfarth et al. 1997). The Salmonella genus comprises more than 2500 serotypes (Popoff 2001), most of which are considered potential human pathogens, but only a reduced number of them have been associated with human infections (Uzzau et al. 2000). In developed countries, outbreaks of non-typhoidal Salmonella infections have been caused mainly by serotypes Enteritidis and Typhimurium (Seyfarth et al. 1997, Davis et al. 1999, Kariuky et al. 1999). S. Typhimurium is a common cause of human salmonellosis in many countries (Tavechio et al. 1996, Leegaard et al. 2000, Esaki et al. 2004, Martinez-Urtaza et al. 2004, Gorman & Adley 2004). It represents the most common serotype isolated from humans and animals in the United States (Bender et al. 2001, Rabatsky et al. 2004) and the second in the United Kingdom (Martinez-Urtaza et al. 2004). Studies reported in the state of São Paulo, Brazil, have also indicated that S. Typhimurium is the second most common serotype isolated from human sources (Tavechio et al.1996) and one of the most frequent from non-human sources (Taunay et al. 1996). The extensive use of antimicrobials in human and veterinary medicine has led to an increase in multidrug resistance strains (MDR) (Threfall et al. 1993, Glynn et al. 1998, Davis et al. 1999, Rabatsky et al. 2004). Fernandes et al. (1992) demonstrated that 95% of S. Typhimurium strains isolated from humans presented high levels of resistance to antimicrobial agents (5-14 marks) and most were isolated in hospital environments, which have been considered the principal vehicle of epidemic MDR S. Typhimurium strains (Riley et al. 1984, Fernandes et al. 1992, Asensi et al. 1995, Kariuky et al. 1999). A MDR S. Typhimurium strain, with definitive phage type 104 (DT 104), has emerged and spread over the world since 1984 (Leegaard et al. 2000, Liebana et al. 2002, Van Duijkeren et al. 2003). The dominant resistance type of these strains is ACSSuT, that is, they are resistant to ampicillin, chloramphenicol, streptomycin, sulfonamides, and tetracycline, and the resistance is encoded by chromosomally located genes (Davis et al. 1999, Izumiya et al. 2001). Studies have shown that only a few phage types tend to dominate within a geographical region for a period of time (Kariuky et al. 1999), and further subtyping of the strains through molecular methods is required in order to detect these epidemiological changes. The aim of this study was to analyse the antimicrobial resistance patterns of S. Typhimurium isolates, received at Institute Adolfo Lutz, São Paulo, Brazil, from 1990-2001, and to investigate the distribution of their phage types, including DT 104, as well as to compare by pulsed-field gel eletrophoresis (PFGE), the genetic heterogeneity of these strains. MATERIALS AND METOHDS Bacterial isolates - A total of 283 S. Typhimurium strains, apparently from sporadic cases of infection, of which 187 from human and 96 from non-human sources, were characterized. These isolates represent 56% of the serotype Typhimurium isolates, from a total of 505, among the 14,249 of Salmonella spp. obtained from the analysis of routine monitoring surveillance, during the period 1990-2001, in the state of São Paulo, Brazil. All the strains were serotyped according to Popoff (2001) at the Laboratory of Enteric Pathogens, Instituto Adolfo Lutz, São Paulo, Brazil. Antimicrobial susceptibility testing - Antimicrobial susceptibility tests were performed by the disk-diffusion method, using disks (Cecon) soaked with amoxicillin-clavulanic acid (30 µg), ampicillin (10 µg), aztreonam (30 µg), cefepime (30 µg), ceftazidime (30 µg), ceftriaxone (30 µg), cephalothin (30 µg), chloramphenicol (30 µg), ciprofloxacin (5 µg), gentamicin (10 µmg), imipenen (10 µg), kanamycin (30 µg), nalidixic acid (30 µg), netilmycin (30 µg), nitrofurantoin (300 µg), sulfamethoxazole-tri-methoprim (25 µg), streptomycin (10 µg), sulfonamides (300 µg), and tetracycline (30 µg). The disk-diffusion method and the interpretation of results were performed as described on the Clinical and Laboratory Standards Institute (CLSI/NCCLS) guidelines. Escherichia coli ATCC 25992 and Pseudomonas aeruginosa ATCC 27583 were used as control on each test (CLSI/NCCLS 2001). MDR was defined as resistance to three or more classes of antimicrobials. Phage typing - Phage typing was performed for all MDR strains and 7% of the susceptible isolates. Phage types were determined by the extended phage typing scheme of Anderson et al. (1977) by using 30 S. Typhi-murium typing phages obtained from the International Reference Laboratory for Enteric Phage Typing, Health Protection Agency, Colindale, London, UK. Phage typing was performed at the National Laboratory for Enteric Pathogens, National Microbiology Laboratory, Health Canada, Winnipeg, Manitoba, Canada, which is the Reference Laboratory for phage typing in the North America, trained by the International Reference Laboratory in Colindale, London, UK. Strains that were untypable through these 30 phages were subtyped with 10 additional experimental phages. All S. Typhimurium strains displaying ununsual phage types, which lyses patterns did not match with the standardized ones, were assigned as atypical (AT or RDNC, reaction does not conform). Such expanded scheme is able to type over than 300 different phage types. PFGE - All MDR strains and 7% of the susceptible isolates were submitted to PFGE for analysis of their heterogeneity as described previously by Thong et al. (1995). Whole cellular DNA of each Salmonella isolated was extracted from a culture of a single colony in 5 ml Luria-Bertani broth for 16-20 (overnight), at 37°C. A volume of 1.5 ml of this culture was transferred into eppendorf tubes and washed twice with 1 ml of cellular buffer (10 mM Tris-HCl, pH 8.0, 1 M NaCl), by centrifuging at 10,000 rpm for 5 min. The pellet was resuspended in 0.2 ml of the same buffer which was kept in a 40°C water bath. This suspension was mixed in equal parts with molten 2% agarose (Bio-Rad Laboratories, Richmond, CA, US) and 22 µl of this mixture was pippeted over a slide surface, forming 10 disks. After solidifying, the disks were transferred to lysis buffer (6 mM Tris-HCl, pH 8.0; 1M NaCl; 100 mM EDTA pH 8.0; 0.5% SDS; 0.5% N-laurilsarcosyl) with 25 µg/ml Rnase A e 1 mg/mg lysozyme, and kept overnight in a 37oC water bath. After incubation overnight at 56°C in 2 ml of lysis buffer with proteinase K (Sigma) at a final concentration of 1 mg/ml, the DNA containing disks were washed five times, for 30 min each, with TE buffer (10 mM Tris-HCl; 1 mM EDTA pH 8.0), on an end to end shaker, and were stored at 4°C in TE buffer. One disk was pre-incubated with 100 µl of 1x restriction buffer (Promega) for 40 min. The buffer was then replaced with a fresh mixture containing 30 U of XbaI restriction enzyme (Promega) in 100 µl fresh restriction buffer and incubated overnight at 37°C. Electrophoresis of the prepared samples was performed on CHEF DR-II system (Bio-Rad Laboratories) by using 1% pulsed-field certified agarose (Sigma), in 0,5 x TBE running buffer (45 mM Tris-HCL; 45 mM boric acid; 1.0 mM EDTA). A lambda ladder pulsed field gel maker (New England Biolabs, Hitchin, UK) was used as a size standard. The electrophoretic conditions were used as follow: Block 1, initial switch time 25 s, final switch time, 65 s, run time 8 h, 6 volts. Block 2, initial switch time 5 s, final switch time 24 s, run time 10 h, 6 volts, temperature 14°C. After electrophoresis the gel was stained for 30 min in sterile distilled water with ethidium bromide (0.5 mg/ml) and distained for 30 min in distilled water and photographed under UV illumination, with Polaroid film. Macrorestriction patterns were compared by using GEL COMPAR II software. Different profiles were designed as A1-An, according to differences in the restriction patterns. A difference of at least one restriction fragment in the patterns was considered the criterion for discriminating (Tennover et al. 1995). RESULTS In a total of 283 strains analyzed, recovered from human and non-human origin during 1990-2001, in different geographic areas of the state of São Paulo, it was shown that 108 (38%) among them presented resistance to at least one antimicrobial agent. The isolates expressed resistance to 11 of the 19 drugs tested, and the highest frequency was observed to tetracyclin (37.5%) followed by streptomycin (24.3%), and sulfonamides (19.4%). Less of 10% of the strains showed resistance (in decrescent order) to ampicillin (9.8%), chloramphenicol (9.5%), sulfamethoxazol-trimethoprim (8.4%), kanamycin (6.3%), nitrofurantoin (6.3%), nalidixic acid (5.6%), gentamicin (2%), and cephalothin (0.7%). None of the isolates were resistant to amoxicillin-clavulanic acid, aztreonam, cefepime, ceftazidime, ceftriaxone, ciprofloxacin, imipenem or netilmycin, and 175 (62%) were susceptible to all antimicrobial agents tested. MDR was identified in 43 (15%) isolates of which 35 (81%) were related with human and eight (19%) with non-human origin. The 23 MDR profiles obtained are shown in Table I, with resistance to three to eight antimicrobials agents. Six strains showed one distinct MDR pattern, ACKSSuTST (ampicillin, chloramphenicol, kanamycin, streptomycin, sulfonamides, sulfamethoxazole-trimethoprim, and tetracycline). Thirty -five strains encompassed 14 DT, and three were UT ones. Eighteen strains were distributed into seven atypical types (RDNC), which did not match to any of the standard phages used for typing. The predominant phage types were DT 193, one of the RDNC patterns and DT 49 with 11, 11 and 6 isolates respectively (Table II). The definitive types 104, 104a, 104b, 104c, or U302 (closely related definitive types, sometimes classified as DT 104 complex) included only three strains in our study: DT 104, DT 104b, and U302. Twenty one PFGE profiles, designed A1-A21, were detected among the 56 strains of S. Typhimurium (Figure) and the most common profiles, were A1 and A8 (Table III). The data in the Figure represent a dendrogram with all PFGE profiles. After cluster analyses of these fingerprints, three different clusters I, II, and III with similarity percentage < 60% were found. In PFGE cluster II, were all isolates of DT 49, belonging to types A1 and A6, DT 193 strains, types A3 and A8 and two strains DT 104 and 104b, type A2. The only one strain of S. Typhimurium U302 was in the cluster I. In cluster III, were the strains of DT193, types A12 and A16, DT 120, type A10, untypables and atypicals ones. The susceptible strains were distributed in clusters I and III. Table III shows the combination of antibiotic resistance, phage types, and PFGE patterns of the MDR studied strains. Type A1 and A6 were the most commum profiles among phage type DT 49 and A3 and A8 among DT 193. Type A2 was found only in DT104 and 104b and A15 in U302. DISCUSSION In this study, we examined a collection of S. Ty-phimurium, isolated from human and non human sources, between 1990 and 2001, in the state of São Paulo, Brazil. Three techniques commonly used for bacterial typing; phage typing, antimicrobial susceptibility testing, and PFGE patterns of XbaI digested cromossomal DNA, were used to characterize these strains, which were epidemiologically unrelated. Antimicrobial susceptibility test demonstrated a high level of drug resistance associated with S. Typhimurium. The data showed that 38% of the strains presented resistance to one to eight antimicrobial agents. The highest frequency of resistance observed in this study was against tetracycline (36.7%), streptomycim (24.3%), sulfonamides (19.4%), ampicillin (9.8%), and chloramphenicol (9.5%), which are also the antimicrobial agents most frequently related to resistance in other parts of the world (Seyfarth et al. 1997, Leegaard et al. 2000, Rabatsky-Ehr et al. 2004). The observation of MDR correlated with studies performed in European Union and United States which confirm that the MDR is common among S. Typhimurium strains, although the frequency found (43 strains, 15%) was lower than in other countries (Leegaard et al. 2000, Nastasi & Mammina 2000, Gorman & Adley 2004, Rabatsky-Ehr et al. 2004). The occurrence of MDR was higher in human (81%) than non-human sources (19%), suggesting that, during this period, 1990-2001, the dissemination of these strains may be attributed to human reservoir and the antimicrobial use in humans has resulted in an increasingly high prevalence of multiple resistance (Fernandes et al. 1992, Asensi et al. 1995, Taunay et al. 1996, Kariuki et al. 1999). All strains of S. Typhimurium studied were susceptible to the second generation fluo-roquinolones, and, 5.6% of them expressed resistance to nalidixic acid (prototypic quinolone). The emergence of nalidixic-resistant Salmonella is a cause of concern, as resistance to fluoroquinolone and reduced susceptibility to fluoroquinolone have been associated with nalidixic acid resistance, since it preceedes resistance to fluo-roquinolone (Hakanen et al. 1999, Gorman & Adley 2004). Although there was no significant predominance of any resistance profile, we have observed in the present study that six MDR strains presented the same mul-tiresistance profile to ampicillin, chloramphenicol, kanamycin, streptomycin, sulfonamides, tetracycline, and sulfamethoxazole-trimethoprim (ACKSSuTTS). They were isolated from human sources in different periods of time and geographical areas and there was no association between phage type, MDR, and PFGE patterns, suggesting that they were not outbreak related and were likely to be from sporadic cases of infection (Table III). Of interest,
in our geographical area link between complex DT 104 (DT104, DT104a,
DT104b, DT104c, and Although in developed countries, only a single phage type has often been associated with outbreaks of MDR S. Typhimurium, our findings did not indicate a significant predominance of any phage type (Table II) and there was no association between phage type and MDR pattern (Table III). We did not also find a significant association between phage type and PFGE pattern, once most phage types were distributed among different PFGE clusters (Table III, Figure). The fact that both strains DT 104 and 104b, presented the same PFGE pattern A2 (Figure) and had been isolated from human source, at the same hospital and time, may suggest a possible link between these strains as a case of infection, either in the community or in the hospital, indicating the importance of the surveillance systems. However, some DT49, DT193, and RDNC strains presented a relationship with PFGE pattern (Table III), it did not suggest any link among them, due to their isolation been done in different time and geographical area. Some strains belonging to these most common phage types DT49, DT 193, and RDNC presented different PFGE patterns (Table III). A similar degree of diversity within DT 193, by PFGE, was revealed in studies described by Doran et al. (2005), indicating that PFGE was able to discriminate between strains close related by phage typing. The strains studied expressed MDR, and belonged to different clusters, presenting different PFGE and phage types, which are widely distributed among hosts and locations and originated from sporadic cases of infection. Kariuki et al. (1999) mentioned similar observations in their studies performed in Nairobi, Kenya, suggesting that these data could be related with developing countries. Continued surveillance of antimicrobial resistance and phage types among S. Typhimurium will monitor the dissemination of MDR strains and outbreaks which can be caused by these potential endemic strains, expressing multiple resistance. ACKNOWLEDGMENTS To Rafiq Ahmed (Health Canada, Winnipeg) who kindly offered training to Ana T Tavechio, on S. Typhimurium phage typing, which was sponsored by the Global Infectious Diseases Research Training Program, directed by Professor Lee Harrison. REFERENCES
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}