 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 101, No. 3, May 2006, pp. 339-340 A micro-spreading improvement for spermatogenic chromosomes from Triatominae (Hemiptera-Reduviidae) Mauricio Camargo/+, María Adelaida Duque-Correa*, Alejandro Berrío Laboratorio de Genética de Poblaciones, SIU, Lab 432 *Laboratorio de Chagas, Instituto de Biología, Universidad de Antioquia, Medellín, Colombia Financial
support: Proyecto de Sostenibilidad Grupo de Ge-nética de
Poblaciones y Mutacarcinogénesis Received
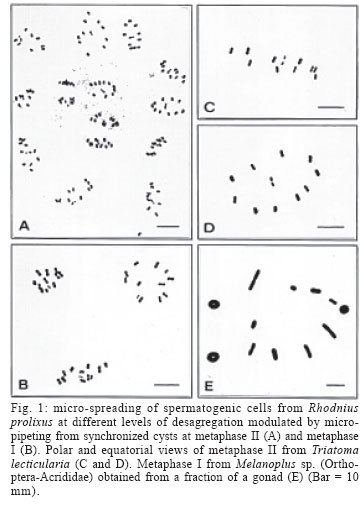
11 November 2005 Code Number: oc06061 Cytogenetics of triatomines have been a valuable biological tool for the study of evolution, taxonomy, and epidemiology of these vectors of Trypanosoma cruzi. Here we present a single microtube protocol that combines micro-centrifugation and micro-spreading, allowing high quality cytogenetic preparations from male gonadal material of Rhodnius prolixus and Triatoma lecticularia. The amount of cellular scattering can be modulated, which can be useful if small aggregates of synchronous cells are desired. Moreover, a higher number of slides per gonad can be obtained with fully flattened clean chromosomal spreads with minimum overlaps, optimal for classical and modern molecular cytogenetic analyses. Key words: micro-spreading - Triatominae - chromosomes The subfamily Triatominae (Hemiptera-Reduviidae) includes 137 species of hematophagous insects, most of them currently recognized as potential vectors of Trypanosoma cruzi, the causative agent of Chagas disease (Galvão et al. 2003). This pathology is recognized as one of the most serious human parasitic diseases of the tropical and subtropical American continent (WHO 2002). The triatomines are mainly distributed in the New World, and occupied diverse habitats from South Argentina to the North American Great Lakes (Crossa et al. 2002). All triatomines so far examined present small holocentric chromosomes, most of them with 20 autosomes and two sex chromosomes (Perez et al. 1992, Panzera et al. 1996). The karyotypic studies are a useful tool utilized to identify the main trends in chromosome evolution, allowing systematic analyses to establish the taxonomic stage of some taxa. Additionally, the cytogenetics of Triatominae has been used to evaluate the cytogeographic relevance of heterochromatic polymorphisms in the epidemiology of this vector related disease (Panzera et al. 1995, 2004, Crossa et al. 2002). Cytogenetics studies in these insects have been traditionally done from gonadal material by squash methods. Here we present an improvement of a micro-centrifugation and micro-spreading protocol, that outdo the overall quality of the cytogenetic preparations, overcoming some of the disadvantages of the classical squash method, and allowing to obtain flattened cells, clean preparations, and consistent chromosomal dispersal, fundamental properties for good cytogenetic and molecular cytogenetic analyses. This modified micro-centrifugation and dropping technique was standardized using male gonads from two triatomine species Rhodnius prolixus and Triatoma lecticularia; internal controls included specimens from Schistocerca sp. and Melanoplus sp. (Orthoptera, Acri-didae). At the end, small amounts (microliters) of material could be handled to produce high quality mitotic and meiotic stages. Adult males from the Triatominae subfamily were obtained from the insectarium of the Chagas Research Group, University of Antioquia at the Sede de Investigaciones Universitarias, Medellín, Colombia. The specimens from the Acrididae family were captured from neighbouring areas of the university campus. The abdomens were opened and the gonads rapidly extracted and immediately immersed in fixative (ethanol:acetic, 3:1), and conserved at 4ºC for days or carry on to the following step. The following process is performed in single microfuge tubes that increase the speed of cell processing, as various centrifugation steps can be performed shortly in a microcentrifuge. Each gonad is immersed in 500 µl of fixative were the first mechanical dispersion step of testicular cells is performed through 8-10 gentle strokes using a P1000 µl micropipette. Cells were pelleted by centrifugation 4 min at 3000 rpm in a microfuge at room temperature (20-22ºC), supernatant discarded with a P200 µl micropipette, and the cell pellet resuspended in 500 µl of fresh fixative. A new thinning-out mechanical step is repeated but with a P200 µl micropipette and 6-10 strokes depending upon the degree of cellular dispersal desired. Cells were pelleted again as described and resuspended in 200 µl of fresh fixative employing a P200 µl micropipette. For slide making, 40 µl of this fine cell suspension is carefully dropped from a distance of 20-30 cm onto 2-3 spots in a detergent-cleaned slides (Extran, Merck). Better results are obtained if the slide is chilled in water-ice and maintains a fine film of water at the time the drops hit the slide. The excess of water-fixative is quickly drained placing the edge of the slide in a paper towel at a short angle (30º), during 5-10 s and/or until the spots of cells in the slide becomes grainy, followed by an immediate heating of the slide from end to end (5-6 s) at a distance of 5-10 cm from the tip of the flame of an alcohol burner. Previous experiences with cultured cells have indicated that this is a critical step for optimal chromosomal spreading, because once the cells touch the surface of the slide, they conserve a spherical shape but flatten gradually as fixative steadily evaporates. Consequently, it is generally observed that the drying rate and length (20-30 s) is more associated with the optimal cellular spreading, and that the dropping height may help to distribute the cells more evenly on the slide surface. Some authors called this period as a narrow time window of chromosome spreading in the process of evaporation of the fixative (Deng et al. 2003). Conventional staining was obtained with Giemsa solution (5%, in phosphate buffer pH 6.8, 12 min). This combination of controlled mechanical scattering, micro-centrifugation and micro-spreading, has several advantages: (a) a high number of slides per gonad can be obtained. In the case of triatomine a minimum of five slide per gonad can be produced if 2-3 micro-drops are used per slide, but could be higher; (b) the amount of cellular dispersal can be modulated, which can be useful if small aggregates of synchronous cells are desired (Fig.1A-B); (c) chromosomal spreads are clean, fully flattened and with minimal overlaps (Fig.1C-E), optimal for classical and molecular cytogenetics analyses such as genomic hybridization and immunofluorescence, well implemented in mammals, but poorly developed in insects; (d) smaller amounts of material can be manipulated and further improvements can be obtained; for example, with Schistocerca and Melanoplus specimens (Acrididae) we have gone down to only one testicular tubuli (Fig. 1E) and it might be possible to micro-centrifuge and micro-spread fractions of it. ACKNOWLEDGEMENTS To the Grupo de Chagas for supplying the Triatominae bugs used in this study. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz |

{kind=link}