 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 101, No. 4, June 2006, pp. 431-435 Anatomical studies of sibling species within Neotropical lymnaeids, snail intermediate hosts of fascioliasis JP Pointier/+, NJ Cazzaniga*, C González-Salas, A Gutiérrez**, JA Arenas***, MD Bargues****, S Mas-Coma**** Ecosystèmes
Aquatiques Tropicaux et Méditerranéens, EPHE, Université,
UMR 5555 du CNRS, 52 Avenue Paul Alduy, Received 7 March
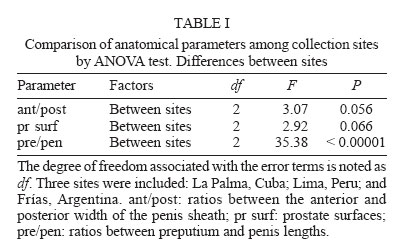
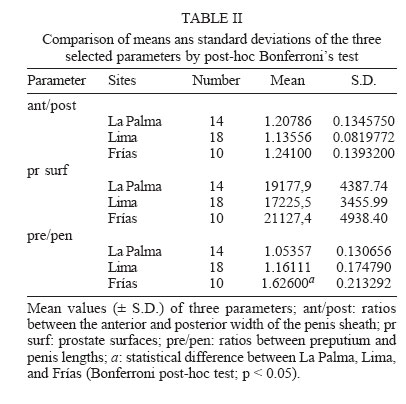
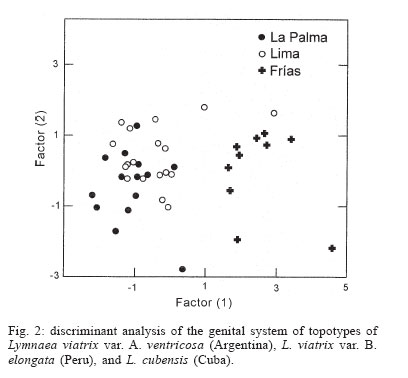
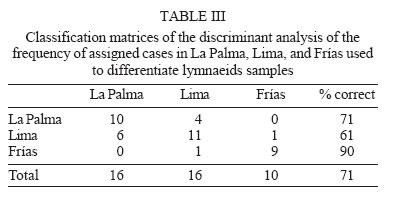
2006 Code Number: oc06077 Several anatomical parameters of the reproductive system have been used to distinguish Lymnaea cubensis from L. viatrix, the snail hosts of fascioliasis in South America and the Caribbean area. Three samples have been collected in the type localities of L. cubensis (Cuba), L. viatrix var. A ventricosa (Argentina, Río Negro Lower Valley), and L. viatrix var. B elongata (Peru, Lima), respectively. Only one parameter, the relative lengths of the penis sheath and preputium, showed significant differences between L. viatrix var. ventricosa and the two other taxa. None of the studied parameters separated L. cubensis from L. viatrix var. elongata. Key words: Neotropical lymnaeids - sibling species - Lymnaea cubensis - Lymnaea viatrix Eleven species of lymnaeid snails have been described in the Caribbean area and South America. However, only six species are considered as valid by Hubendick (1951) and Paraense (1976, 1982a, 1983, 1984, 1986, 1994, 1995): Lymnaea viatrix d'Orbigny, 1835; L. diaphana King, 1830; L. cubensis Pfeiffer, 1839; L. cousini Jousseaume, 1887; L. rupestris Paraense, 1982 and L. columella Say, 1817. This last species has now a very large distribution in all tropical countries particularly in the neotropical area (see Malek & Chrosciechowski 1964, Paraense 1982b, Gomez et al. 1986). Two other species may be added to that list (i) L. truncatula (Müller, 1774) from Europe which recently invaded several Andean countries (Oviedo et al. 1995, Jabbour-Zahab et al. 1997, Mas-Coma et al. 2001, Durand et al. 2002, Bargues & Mas-Coma 2005); (ii) L. plicata Hylton-Scott, 1953 from Chubut Province, Argentina which validity has not yet been confirmed. The morphology of the shell and reproductive system of five of those species has been well studied and allows a clear identification (Paraense 1982a, 1984, 1986, 1995, Samadi et al. 2000, Pointier et al. 2004). However, the case of two taxa, L. viatrix and L. cubensis, remains unclear. Regarding L. viatrix, d'Orbigny (1835) described two varieties (i) var. A ventricosa from the Río Negro Lower Valley, Patagonia, Argentina, and (ii) var. B elongata from Lima, Peru. A first study carried out by Paraense (1976) on topotypic specimens from Argentina and Peru found them anatomically identical. In a subsequent paper, this author also considered L. cubensis from Soroa, Cuba, anatomically indistinguishable from L. viatrix (Paraense 1982b). Moreover, an analysis of the genetic variability based on 12 enzyme loci revealed different fixed alleles at nine loci between two syntopic samples of L. viatrix var. elongata at the type locality in Lima, Peru (Durand et al. 2002). The absence of heterozygotes within this locality thus indicated the presence of isolated populations or cryptic species within the variety elongata. In the same study, Durand et al. (2002) also found significant genetic differences between the two taxa from Lima and another population sampled in Bahia Blanca, Argentina, located about 350 km to the north of the type locality for L. viatrix var. ventricosa. More recently, the use of several molecular markers (PCR-RLFP analysis of the ITS1 and ITS2 rDNA and 16S rDNAmt using 12 restriction enzymes) in a study of several populations morphologically identified as L. viatrix from Brazil, Uruguay, and Argentina showed a high level of genetic polymorphism between populations which was interpreted as an intra-specific variation (Carvalho et al. 2004). A morphological comparison of several populations of L. cubensis from the Caribbean area and several populations of the introduced L. truncatula from Bolivia was carried out by Samadi et al. (2000) using several con-chological and anatomical characters of the reproductive system. The conchological study showed the presence of a large amount of variability within and between populations that did not allow distinction between taxa. The analysis of anatomical parameters of the reproductive system also showed a high variability, but all these characters clearly separated L. cubensis from L. truncatula independently from their geographical origin. In the present paper, we use these anatomical characters in the same way to see if it is possible to distinguish morphologically L. cubensis from L. viatrix. The material studied included samples from the type localities of L. cubensis (Cuba), L. viatrix var. A ventricosa (Argentina, Río Negro Lower Valley), and L. viatrix var. B elongata (Peru, Lima). MATERIALS AND METHODS Snails from three populations were sampled in the type localities of L. cubensis, L. viatrix, var. A ventricosa, and L. viatrix, var. B elongata, respectively. Regarding the type locality of L. cubensis, no specific region of Cuba was indicated by Pfeiffer (1839). A sample of 32 snails was collected in La Palma area, Cuba (N 22° 45'; W 83° 34'). The original locality of L. viatrix var. ventricosa was described by d'Orbigny as the banks of the Río Negro as far as 30-40 km before its mouth. In 1973, Paraense collected a sample from irrigation ditches at the city of Viedma (S 40° 49'; W 63° 00'), a place within the area of Patagonia indicated by d'Orbigny as the type locality for this snail (Paraense 1976). In 2004, these ditches did not exist any more and careful searches in this area resulted in the discovery of a single lymnaeid population in a locality named Teniente General Frías (S 40° 14' 98; W 64° 10' 85) on the Río Negro banks about 120 km to the west of Viedma. A total of 36 snails was sampled at this site. The Río Negro Lower Valley is a geographical space that centers on an ecologically homogeneous oasis subregion to the north of the Somuncurá plateau. Climate, physiography, soils, and vegetation are quite uniform from Choele Choel (S 39° 17', W 65° 35', 132 m above sea level) towards the east, through to the mouth of the Río Negro River. Annual rainfall is 230-325 mm, while potential evapotranspiration is 600-680 mm, so the hydric balance is always negative (Cuevas-Acevedo 1981). The original locality of L. viatrix var. elongata was described by d'Orbigny as the surroundings of Lima, Peru. A total of 65 snails was thus collected along the banks of Río Lurin in the suburbs of Lima, Peru (S 12° 3'; W 77° 4'). Snails collected in the field were allowed to relax overnight using menthol. They were then immersed for 40 s in hot water (70°C), from which they were transferred to water at room temperature. The soft parts (under water) were drawn from the shells with forceps applied to the cephalopedal mass, and fixed in slightly modified Railliet-Henry's fluid (distilled water 930 ml, sodium chloride 6 g, 40% formalin 50 ml, glacial acetic acid 20 ml). In order to compare the three samples without size/age bias, only adult snails of a similar size (6.5 to 7.8 mm) were used for anatomical studies. Snails were dissected under a stereoscopic microscope and drawings of the reproductive system were made using a camera lucida attachment. The pictures obtained were digitalized with an `Apple Scanner'. The prostate surfaces (pr surf) were then measured on 18 specimens from Lima, Peru, 14 specimens from La Palma, Cuba and 10 specimens from Frías, Argentina. In addition, two other characters were measured: the ratio between preputium and penis lengths (pre/pen), and the ratio between the anterior and posterior width of the penis sheath (ant/post; Fig. 1). Anatomical data were tested for normality and homogeneity of variances using Shapiro-Wilks W statistics and Bartlett's test respectively. The anatomical parameters were then compared using an analysis of variance (ANOVA, Sokal & Rohlf 1995) for the three sites. When differences were detected by ANOVA, Bonferroni' tests were used since the number of snails at each site were not the same. Canonical discriminant-function analyses (Blackith & Reyment 1971) were also used in support to the ANOVA to determine differences between sites. For this last analysis we used a data array of `pre/pen', `ant/post', and `pr surf' as variables. Thus, each site was described by three variables. RESULTS Whatever anatomical parameter studied, no departures from normality and homogeneity of variances were found. The ANOVA results showed significant differences between sites for only one anatomical character: the pre/pen ratio (Table I). Multiple range test showed that the significant differences were found between La Palma and Frías and between Lima and Frías. No difference was observed between La Palma and Lima (Table II). Discriminant analysis showed differences between sites (Fig. 2). The first eigenvalue (92.1%) is very large relative to the second (7.89%), indicating that the first axis captures most of the variance among sites. These two canonical discriminant functions account for more than 99% of the total dispersion of the groups. The classification success of the discriminant analysis (Table III) indicates a high separation between Frías and the two other sites (Wilks' statistic l = 0.3048; p < 0.0001). Overall, 71% of individuals were correctly classified (Table III). DISCUSSION Conchological studies of lymnaeid snails carried out by Samadi et al. (2000) showed the presence of a large amount of variability that did not allow distinction between L. truncatula samples from the Old World and Bolivia on the one hand and L. cubensis samples from the Caribbean area on the other hand. However, those authors observed a clear anatomical distinction between the two taxa using charaters of the male reproductive system. All the selected anatomical parameters actually separated the two groups of samples, and confirmed the two genetic clusters that were previously obtained using allozyme electrophoresis (Jabbour-Zahab et al. 1997). The most relevant anatomical characters were the size of the prostate and the relative lengths of the penis sheath and preputium. The choice of these parameters in our analysis on topotypic specimens of L. cubensis and L. viatrix also showed a great variability and only one parameter, the relative lengths of the preputium and penis sheath, allowed the distinction between L. viatrix var. ventricosa on the one hand and L. viatrix var elongata and L. cubensis on the other hand. This statement is supported by the high classification success of the discriminant analysis carried out on these samples. None of the studied parameters allowed a significant separation between the viatrix var. elongata and cubensis samples. However, it is noteworthy that the analysis of the genetic diversity of the same samples has shown that L. viatrix var. elongata and L. cubensis are quite distinct genetic entities (Durand et al. 2002). These results point out the difficulty of correctly identifying the species belonging to this group of lymnaeids using only anatomical characters and confirm the previous studies carried out by Paraense (1976, 1982b). The relative lengths of the penis sheath and preputium appear as a relevant character, which could be used to differentiate some taxa but our study also showed that this character is of limited value for this group of lymnaeids. Further morphological and molecular studies carried out on much more populations covering the whole distribution area of these lymnaeids are necessary to a better understanding of the taxonomy of this group. Lymnaeid snails identified as viatrix or cubensis are commonly cited in the literature as playing the role of intermediate host for the liver fluke Fasciola hepatica (Morales & Pino 1981, Pino & Morales 1982, Lara et al. 1988, Larrea et al 1993, Rangel-Ruiz 1993, Kleiman et al. 2004, Mas-Coma et al. 2005). Unfortunately, due to the difficulty of a clear morphological identification, it is quite impossible to know which species is actually involved in the transmission of the parasite in order to develop effective measures of control. For example, Ueno et al (1975) reported the occurrence of two intermediate hosts of F. hepatica in the Bolivian Altiplano, L. viatrix, and L. cubensis. As a matter of fact, subsequent studies showed the role of a single species, the introduced L. truncatula (Oviedo et al. 1995, Jabbour-Zahab et al. 1997, Mas-Coma et al.1999, Mas-Coma et al. 2001, Bargues & Mas-Coma 2005). Moreover, molecular studies using microsatellite markers revealed the presence of a single genotype of L. truncatula in the Bolivian Altiplano (Meunier et al. 2001). This discovery has important epidemiological consequences because the occurrence of a single species with an absolute lack of genetic variability could greatly facilitate its control. ACKNOWLEDGEMENTS To Dr J Boissier for helpful revision of the manuscript. REFERENCES
The following images related to this document are available:Photo images[oc06077t2.jpg] [oc06077f2.jpg] [oc06077t1.jpg] [oc06077t3.jpg] [oc06077f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}