 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 101, No. 4, June 2006, pp. 455-457 Molecular characterization of human Trypanosoma cruzi isolates from endemic areas in Panama Octavio E Sousa/+, Franklyn Samudio*, Corina de Juncá, José E Calzada* Center for Research
and Diagnosis of Parasitic Diseases, Faculty of Medicine, University of Panama,
Panama *Instituto Conmemorativo Gorgas de Estudios de la Salud, Panama, Panama Financial partial support: grant PAN 6010, International Atomic Energy Agency Received 5 January
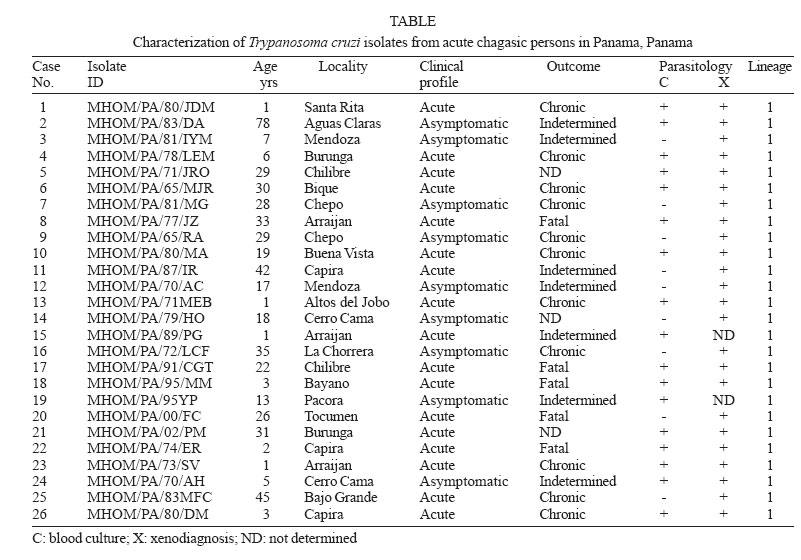
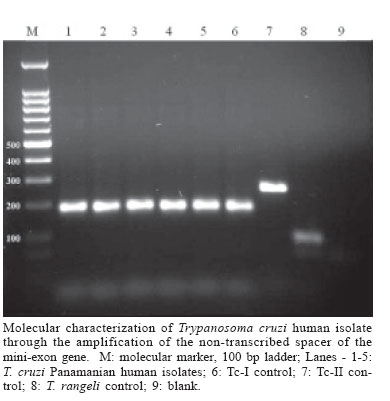
2006 Code Number: oc06080 The present work provides information on Trypanosoma cruzi genotype circulating in endemic areas of Chagas disease in Panama. A total of 26 crude stocks of T. cruzi, isolated from the blood of persons with different clinical profiles of Chagas disease were collected and crio-conserved until used. Most of the stocks had been characterized by means of isoenzyme electrophoresis on cellulose acetate membranes. The clinical profiles of infected persons included 9 (34.6%) asymptomatic and 17 acute (65.4%) including 5 (19.2%) fatal cases, 2 under 5 years old and 3 adults. A multiplex-PCR assay based on the amplification of the non-transcribed spacer of the mini-exon gene was performed. All stocks of T. cruzi included in the study were found to correspond to Tc I group. This result supports the predominance of T. cruzi-I in the transmission cycles affecting the human population in the Republic of Panama. Key words: Trypanosoma cruzi - lineage - acute Chagas disease - Panama Trypanosoma cruzi is a blood parasite of man in America, often resulting in clinical manifestations and pathologies of Chagas disease. It is generally accepted that some 18 million people have been demonstrated to be infected by this parasite. Chagas disease, first described by Carlos Chagas in 1909 from endemic areas of Brazil, is naturally recognized as a zoonosis that affects the human population of endemic areas of Latin America, from Argentina and Chile in the south, to Mexico and Southern United States in the north. Transmission of the parasite results from contamination with the infective trypanosomes found in the feces of naturally infected triatomine vectors that exist in sylvatic, rural or domiciliary habitats. Chagas disease has been recorded in the Republic of Panama since 1930, when trypomastigotes of T. cruzi were found in the blood of three hospitalized children suspected of malarial fever. A report of these cases was published by Miller (1931). Chagas disease, in Panama, is more frequently reported from rural areas on both sides of the Panama Canal (Sousa & Johnson 1971). In these areas, the parasite is primarily transmitted by Rhodnius pallescens, a triatomine vector found in close association with the palm-tree (Attalea butyracea) in domestic, peridomiciliary, and sylvatic ecotopes (Whitlaw & Chaniotis 1978). The infection in man can result in asymptomatic, clinical, and severe, at times fatal disease. However, when symptomatic the disease in Panama generally runs a milder course than in T. cruzi infections acquired in endemic areas of the Southern Cone of America (Nuñez 1966). In Panama the infection is primarily of the cardiotropic form during the acute and chronic phases with no evidence of the digestive form developing during later stages of the infection. Since the early pathological studies there is no evidence of chagasic megasyndromes (Blandon et al. 1969). The situation appears to be similar to that of Venezuela (Añez et al. 2004), Amazonia, Brazil (Coura et al. 1999), and part of Central America (Ruiz-Sanchez et al. 2005). Studies on the genetic variability of T. cruzi have recently demonstrated two primary phylogenetic lineages, Tc-I and Tc-II, with a third group of hybrid strains still under revision. Further studies identified and supported six T. cruzi lineages (Brisse et al. 2000) including T.cruzi-I and T.cruzi-II. The high polymorphism of T. cruzi and the significance of its genetic variability in the pathogenesis of Chagas disease has been considered (Macedo et al. 2004) and led to establish the basis for a clonal-histotropic model for Chagas disease (Macedo et al. 2002). Geographical distribution and epidemiological considerations such as natural sources of infections, vectors and reservoirs, have been studied resulting in several suggested associations (Miles et al. 2005, Yeo et al. 2005). T. cruzi-I, appears associated with American marsupials, Didelphis spp., arboreal ecologies and northern distribution, while T. cruzi-II are more associated with armadillos, terrestrial ecologies and Southern-Cone areas in America. The parasites analyzed were obtained from the blood of patients during the acute phase of the disease; people with parasitaemia demonstrated through hemoculture and/or xenodiagnosis, including asymptomatic, clinical, and fatal cases. This study provides information on the genotype of T. cruzi affecting man in this area, in an effort to establish their relationship to the different groups known. A total of 26 crude stocks of T. cruzi, isolated from the blood of patients with different clinical profiles of Chagas disease were collected and maintained in liquid nitrogen. Most of the stocks had been characterized by means of isoenzyme electrophoresis on cellulose acetate membranes as previously described (Kreutzer & Sousa 1981). Information regarding the case, identification of the stock, location, and year of isolation is presented in the Table. Parasites were grown in liver infusion tryptose medium (LIT) supplemented with 10% bovine fetal serum until logarithmic phase was reached. Parasites were collected by centrifugation (2500 g at 4°C for 10 min) and washed twice with buffer PBS (pH 7.2). The pellet was used for DNA extraction using a commercial kit following manufacturer' instructions (Wizard Genomic DNA Purification System, Promega, US). DNA integrity was checked in 1% agarose gels and DNA concentration estimated by spectrophotometry at 260 nm. A multiplex-PCR assay based on the amplification of the non-transcribed spacer of the mini-exon gene (Fernandez et al. 2001) was performed on the Panamanian T. cruzi stocks using 25 ng of DNA. The results are summarized in the Table, including CL-Brener and Y strains as Tc-II controls, as well as a local T. rangeli isolate from Panama. All stocks of T. cruzi included in the study amplified a 200 bp product which corresponds to group 1 (Figure). No evidence of TcII was found. The clinical profiles of infected patients included 9 (34.6%) asymptomatic, and 17 acute (65.4%) including 5 (19.2%) fatal cases, 2 under 5 years and 3 adults. It has been our experience that in this area it is extremely difficult to isolate parasites through blood culture or serial xenodiagnosis during the chronic stage of Chagas disease (Sousa 1988). Six patients included in the present work were periodically examined during a prolonged period (11 to 23 years) after the initial parasitemia without evidence of parasites through blood cultures or xenodiagnosis but, remaining seropositive against T. cruzi. In Panama there appears to exist a single lineage involved in the epidemiological factors of transmission in the rural endemic area of Chagas disease. T. cruzi-I predominates in this cycle involving R. pallescens as vector, the palm-tree A. butyracea the ecotope, and Didelphis marsupialis as primary reservoir. The present work is further supported by studies on 60 T. cruzi stocks found in naturally infected R. pallescens collected in 30 royal palm trees (A. butyracea) from three different localities from endemic areas of Chagas disease in Panama (data not shown) as well as parasites obtained from the blood of a domestic dog and one common opossum (D. marsupialis). All stocks tested have been characterized as T. cruzi-I. It is not yet clear whether clonal T. cruzi-II may be present in reservoir hosts or vectors other than R. pallescens. The situation in this country is similar to that described for the Amazon region of Brasil (Coura et al.1999), Colombia (Cuervo et al. 2002), Venezuela (Añez et al. 2004), and Guatemala and Mexico (Ruiz-Sanchez et al. 2005) where T. cruzi-I predominates among the parasites recovered from endemic areas of the disease. This condition contrasts with the asumption that Chagas disease is due to a single parasite lineage, T. cruzi II and argues against human infection cases exclusively due to T. cruzi I strains (Di Noia et al. 2002). REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06080t1.jpg] [oc06080f1.jpg] |
| |||||||||

{kind=link}
{kind=link}