 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 101, No. 5, August, 2006, pp. 511-516 Comparison between indirect enzyme-linked immunosorbent assays for Anaplasma marginale antibodies with recombinant major surface protein 5 and initial body antigens Virgínia MG Silva*, Flábio R Araújo, Claudio R Madruga/+, Cleber O Soares, Raul H Kessler, Maria AO Almeida**, Stênio P Fragoso***, Lenita R Santos*, Carlos AN Ramos, Gisele Bacanelli, Roberto AA Torres Júnior Embrapa Gado de
Corte, Caixa Postal 154, 79002-970 Campo Grande, MS, Brasil *Pós-Graduação
em Imunologia, Instituto de Ciências da Saúde **Departamento
de Patologia e Clínicas, Escola de Medicina Veterinária, Universidade
Federal da Bahia, Salvador, BA, Brasil ***Instituto de Biologia Molecular
do Paraná, Curitiba, PR, Brasil Financial support: Fundect, CNPq Received 25 November
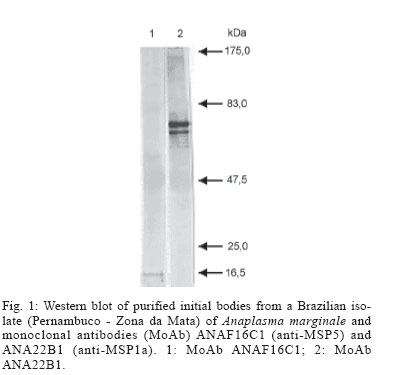
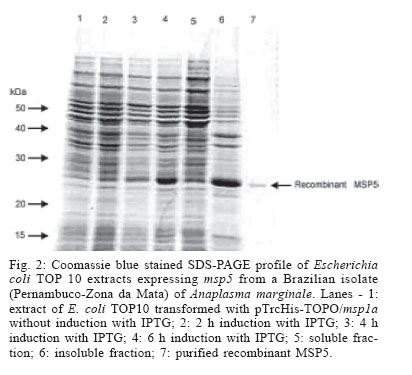
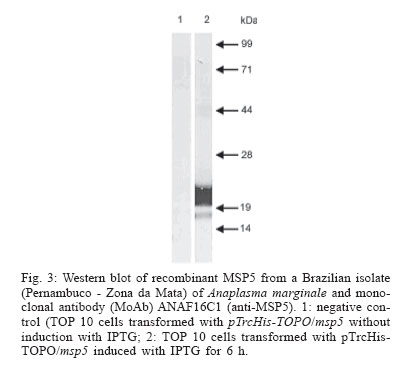
2005 Code Number: oc06086 Indirect enzyme-linked immunosorbent assays (ELISAs) based on recombinant major surface protein 5 (rMSP5) and initial body (IB) antigens from a Brazilian isolate of Anaplasma marginale were developed to detect antibodies against this rickettsia in cattle. Both tests showed the same sensitivity (98.2%) and specificities (100% for rMSP5 and 93.8% for IB ELISA) which did not differ statistically. No cross-reactions were detected with Babesia bigemina antibodies, but 5 (rMSP5 ELISA) to 15% (IB ELISA) of cross-reactions were detected with B. bovis antibodies. However, such difference was not statistically significant. Prevalences of seropositive crossbred beef cattle raised extensively in Miranda county, state of Mato Grosso do Sul, Brazil, were 78.1% by rMSP5 ELISA and 79.7% by IB ELISA. In the analysis of sera from dairy calves naturally-infected with A. marginale, the dynamics of antibody production was very similar between both tests, with maternal antibodies reaching the lowest levels at 15-30 days, followed by an increase in the mean optical densities in both ELISAs, suggesting the development of active immunity against A. marginale. Results showed that all calves were seropositive by one-year old, characterizing a situation of enzootic stability. The similar performances of the ELISAs suggest that both tests can be used in epidemiological surveys for detection of antibodies to A. marginale in cattle. Key words: Anaplasma marginale - enzyme-linked immunosorbent assay - major surface protein 5 - initial body - Brazil Anaplasmosis is an important vector-borne hemopara-sitic rickettsial disease of ruminant livestock in tropical and subtropical regions of the world (Palmer et al. 1986), including South America (Barros et al. 2005). The disease is caused by Anaplasma marginale, an ehrlichial pathogen of the genogroup II (Dumler et al. 2001), and is characterized by fever, anemia, general weakness, pallor mucous membranes, weight loss (Ajayi et al. 1978), abortion (Correa et al. 1978), decreased milk production, and mortality during the acute phase of infection (Palmer et al. 1986). Control of A. marginale infection requires both an effective vaccine and accurate identification of carrier cattle (Andrade et al. 2004). Two possible methods for routine carrier detection are identification of rickettsial DNA in blood and the detection of A. marginale-specific antibodies in serum (McGuire et al. 1991, Knowles et al. 1996). Despite the high sensitivity and specificity of the former ones, serological methods remain the most wildly used diagnostic tools. Serologic tests for anaplasmosis based on crude antigen have a high error rate due to false-positive results with bovine erythrocyte proteins contaminating the A. marginale antigen and reacting with anti-erythrocyte antibodies in bovine sera (as reviewed by Knowles et al. 1996). To improve serological diagnosis of bovine anaplasmosis, research has focused on the identification and characterization of A. marginale membrane antigens, by gene cloning and production of recombinant proteins, which would be suitable for diagnostic tests (Reyna-Bello et al. 1998). Six major surface proteins (MSPs) have been initially described: MSP1a, MSP1b, MSP2, MSP3, MSP4, and MSP5 (Palmer et al. 1986, Oberle et al. 1988, Visser et al. 1992). Recently, four new membrane proteins were identified: Ana29, Ana 32, Ana 37, and Ana 43 (Riding et al. 2003). From these, the immunodominant 19-kDa MSP5 has potential use in serological tests (Ndung'u et al. 1995, Knowles et al. 1996, Reyna-Bello et al. 1998, Torioni de Echaide 1998), as it is conserved between Anaplasma species and isolates, including Brazilian isolates of A. marginale (Visser et al. 1992, Kano et al. 2002, Oliveira et al. 2003). In this study, we have compared recombinant MSP5 and initial body proteins as ELISA antigens for Anaplasma antibodies in Brazil. MATERIALS AND METHODS A. marginale isolate - Initial bodies (IB) were isolated from erythrocytes from a splenectomized calf experimentally infected with a Brazilian strain of A. marginale (Pernambuco, Zona da Mata). Blood was collected in vacuum tubes with heparin when rickettsemia reached 87%. Purification of IB - A. marginale IB were isolated from infected erythrocytes by differential centrifugation, as described by Palmer and McGuire (1984). Purification was certified by comparison in the sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) with non-infected bovine erythrocytes. Western blots with monoclonal antibodies (MoAbs) ANAF16C1 and ANA22B1, to MSP5 and MSP1a, respectively (Visser et al. 1992, Palmer et al. 1988), were also performed, as described elsewhere (Oliveira et al. 2003). Genomic DNA extraction - DNA was purified from 300 µl of infected blood by Easy DNA kit (Invitrogen, US). The concentration of extracted DNA and the A260/A280 ratio were determined on a GeneQuant spectrophotometer (Amersham Pharmacia Biotech, US). Amplification of msp5 Primers were designed to amplify a 714 bp fragment, including the entire msp5 gene and a 3' UTR fragment, according to the reported msp5 nucleotide sequence (Genbank accession no. M93392). These primers were msp5F (5'-ATGAGAATTTTCAA GATTGTGTCTAACCTT-3') and msp5R (5'-AGGAAAG CCCCCAAAGCCCCATACTT-3'). PCR amplification reaction mixture was prepared in a volume of 50 µl, containing 10 mM Tris-HCl (pH 8.5), 50 mM KCl, 1.5 mM MgCl2, 0.2 mM (each) deoxynucleoside triphosphate, 100 hg (each) primer, 100 hg of genomic DNA, and 2.5 U of Taq DNA polymerase (Cenbiot, Brazil). Amplification was performed in a PTC-200 thermocycler (MJ Research, US) as follows: 94°C for 1 min (denaturation); the next 35 cycles were 94°C for 1 min (denaturation), 60°C for 1 min (annealing), and 72°C for 30 sec (extension), and the last cycle was 72°C for 7 min (extension). PCR products were analyzed by electrophoresis in 1% agarose gel, stained with ethidium bromide (0.5 mg/ml). Cloning of msp5 - PCR amplicon was cloned in plasmid pTrcHisTOPO (Invitrogen), according to the manu-facturer's instructions. The recombinant plasmid was used to transform Escherichia coli TOP10 cells. Insert orientation was determined by colony PCR, using plasmid primer Xpress (Invitrogen) and primer msp5R. The insert was also sequenced in both directions by the dideoxide method and analyzed in an ABI 3100 automatic sequencer (Applied Byosystems, US). Only sequences with a PHRED index greater than 20 were considered. Recombinant MSP5 production - An overnight 50 ml culture of E. coli TOP10 carrying the recombinant plasmid grown in liquid selective Luria-Bertani (LB) medium containing 100 µg/ml of ampicillin (LB-amp) was cultured in 450 ml of LB-amp for 1 h at 37°C. Isopropyl-β -D-thiogalactopyranoside (IPTG) was added at a final concentration of 1 mM. The cultures were allowed to grow for 6 h. Aliquots of 2 ml were taken before induction with IPTG and then every 2 h. These aliquots were centrifuged at 18,000 x g and the pellet was frozen at - 20°C. After 6 h culture, E. coli cells were recovered by centrifugation at 10,000 x g for 10 min. The pellet was suspended in 20 ml of 50 mM phosphate buffer and then frozen in liquid nitrogen. After thawing at 4ºC, the suspension was sonicated (Sonifier 250, Branson, US - output 40) for 6 x 20 s on ice. The recombinant protein production was verified by SDS-PAGE, followed by Coomassie blue staining. The identity of rMSP5 was confirmed by Western blot with monoclonal antibody ANAF16C1, as described elsewhere (Oliveira et al. 2003). Purification of MSP5 The cell lysate was homogenized with 30 ml of 50 mM phosphate buffer and 50 ml of 2 x SDS-PAGE sample buffer. The material was boiled for 5 min and submitted to electrophoresis in a 2 mm preparative 15% SDS-polyacrilamide gel (Hoeffer 600). After the protein separation, the gel was immersed on cold 1 M potassium acetate (Neophytou et al. 1996). The major protein band was gel excised and electro dialyzed in glycine buffer at 35 mA, for 3 h, at 4ºC. The protein was dialyzed overnight against 5 M urea, 1 mM tris-HCl, pH 7.7, and 1% 2-mercaptoethanol, at 35 mA for SDS removal (Tuszynski & Warren 1975), and dialyzed overnight against 50 mM phosphate buffer, at 4ºC, for urea removal. Purity of the recombinant protein was assessed by SDS-PAGE. ELISA standardization - optimal dilutions of antigen, sera and conjugate for ELISAs with rMSP5 and with IB were determined by testing 16 sera negative for A. marginale antibodies, from cattle kept in an isolation area of Embrapa Beef Cattle Research Center, and 16 sera positive for A. marginale antibodies, from experimentally-infected cattle. ELISA with IB antigen - 96-well plates (ref. 3590, Costar, US) were adsorbed with 350 hg/well of IB proteins, diluted in adsorption buffer (1M Na2HPO4.2H2O, 0.8 M C6H6O7.H2O, pH 4.0), for 12 h, at 4ºC. Plates were then kept at - 20ºC. After thawing, the plates were washed five times with phosphate-buffered saline (PBS) with 0.1% tween (PBST) and were blocked with Dulbelcco's buffered saline, pH 7.2, with 2% γ-globulin free equine serum and 5% slim milk, for 4 h, at 4ºC. After five washes with PBST, positive and negative sera (in quadruplicate) and test sera (in duplicate), diluted 1:1,000 in PBST, were incubated for 30 min at 37ºC. Plates were then washed five times with PBST, and 100 µl/well of rabbit anti-bovine IgG horseradish peroxidase conjugate (ref. A-7414, Sigma, US), diluted 1:5000 in PBST, were added. The plates were incubated as described and after 10 washes with PBST, the chromogen/substrate o-phenylene diamine dihydrochlo-ride (OPD; Sigma)/ H2O2 was added. The reaction was stopped by addition of H2SO4 (2.5 N) and results were read on an El x 800 ELISA reader (Bio-Tek, US), with a filter of 490 ηm. ELISA with rMSP5 - 96-well plates (ref. 3590, Costar) were adsorbed with 3 hg/well of rMSP5, diluted in Dulbelcco's phosphate buffer, pH 7.2, for 12 h, at 4ºC. Plates were then kept at - 20ºC. After thawing, plates were blocked with PBST with 2% γ-globulin free equine serum and 5% slim milk, for 30 min, at 37ºC. After five washes with PBST, positive and negative sera (in quadruplicate) and test sera (in duplicate), diluted 1:50 in PBST, were incubated for 40 min at 37ºC. Plates were then washed five times with PBST, and 100 µl/well of rabbit anti-bovine IgG horseradish peroxidase conjugate (ref. A-7414, Sigma), diluted 1:10,000 in PBST, were added. Plates were incubated for 30 min at 37ºC and after 10 washes with PBST, the chromogen/substrate substrate o-phenylene diamine dihydrochloride (OPD; Sigma)/ H2O2 was added. The reaction was stopped by addition of H2SO4 (2,5 N) and results were read on a ELx-800 ELISA reader (Bio-Tek), with a 490 hm filter. Cut-off determination - Cut-offs were determined as the mean optical density (OD) of 20 negative control sera plus three standard deviations. Determination of sensitivity and specificity of the tests - Sensitivity of both tests was determined with 57 cattle sera positive for A. marginale antibodies by immunofluorescent antibody test (IFAT). Specificity was determined with 65 cattle sera negative for A. marginale antibodies by IFAT. Cross-reactions with Babesia antibodies - Cross-reactions with Babesia antibodies were evaluated in 20 cattle sera positive for B. bovis or B. bigemina antibodies and negative for A. marginale antibodies by IFAT. Prevalence of A. marginale in cattle raised extensively in Mato Grosso do Sul - Sera were collected from 128 cross-bred cows (predominantly Nelore) of a transition area between Brazilian Savanna and Pantanal regions, in the county of Miranda, Mato Grosso do Sul, according to the statistical model of the Centro Pan-Americano de Zoonoses (1979), that considered 95% of confidence degree and 2% of error probability. Cattle were raised extensively and ticks were not controlled in these farms, due to the low infestations in beef cattle. Sera from these calves were analyzed by both ELISAs for prevalence determination and comparison. Evaluation of the ELISAs with sera from calves naturally infected with A. marginale - Sera from 28 Holstein calves from a dairy farm, located on an endemic region for anaplasmosis (São Pedro county, São Paulo, Brazil) were collected from 15 days up to one year of age and analyzed by IB ELISA and rMSP5 ELISA. After birth, these calves remained with their mothers for 24 h, and then were kept in individual stalls until they reached four months. In the individual stalls, calves received four liters of colostrum for two days and then four liters of milk and ration ad libitum. After four months, calves were kept in collective pastures, where they were fed with silage, hay and ration. Statistical analysis - Data were analyzed by Qui-square test or Fischer's exact test, with a confidence interval of 95%. RESULTS IB antigen production - The IB were partially purified from infected erythrocytes. In the SDS-PAGE analysis, four erythrocyte bands were detected in the IB antigen (data not shown). In the Western blot, monoclonal antibodies ANAF16C1 (MSP5) and ANA22B1 (MSP1a) reacted with epitopes of 16 kDa and 80 kDa, respectively (Fig. 1). Amplification and cloning of msp5 - As expected, a 714 bp fragment was amplified with primers msp5F and msp5R, which was successfully cloned in pTrcHis-TOPO. It was possible to sequence a 633 bp fragment of the insert, which corresponds to the coding sequence of MSP5. The resulting DNA sequence (Genbank accession no. AY245428) showed 98.4% of identity (623/633 nucleotides) with msp5 from A. marginale Florida isolate (Genbank accession no. M93392) in the BLASTn analysis. At the protein level, the similarity between isolates was 99%. Expression of rMSP5 - A 22 kDa protein was expressed (Fig. 2), which reacted with MoAb ANAF16C1 in the Western blot with cell lysate (Fig. 3). No reaction was detected with non-induced transformed TOP10 cells. Performances
of rMSP5 and IB ELISAs After the initial standardization
of rMSP5 and IB ELISAS, the performances of both tests were evaluated.
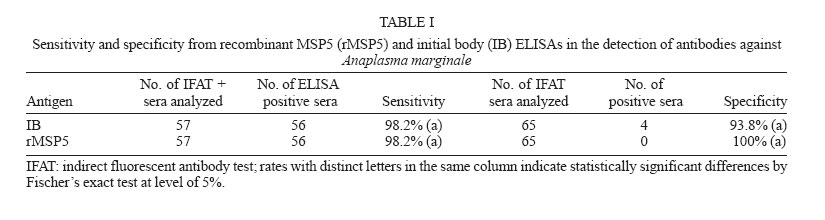
The analysis of sensitivity and specificity from both tests in shown in Table
I. There were no statistical differences neither in the sensitivities
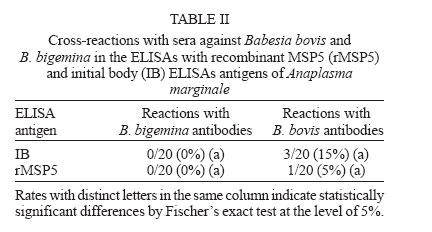
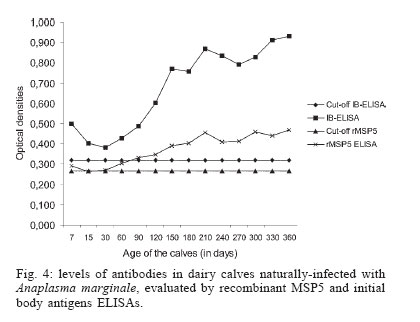
nor in the specificities between both tests Cross-reactions with Babesia sp. antibodies - No cross-reactions with B. bigemina antibodies were verified in both ELISAs. From the 20 serum samples positive for B. bovis antibodies in the IFAT, three (15%) reacted with IB antigen and one (5%) reacted with rMSP5 (Table II). Differences between both tests were not statistically significant (P > 0.05). Prevalence of A. marginale antibodies in an extensively-raised cattle herd - Prevalences of A. marginale antibodies in an extensively-raised cattle in Miranda county, Mato Grosso do Sul, by IB and rMSP5 ELISAs were 79.7 and 78.1%, respectively (Table III). There were no statistical differences between both tests (P > 0.05). Evaluation of the ELISAs with sera from calves naturally-infected with A. marginale - The analysis of sera from 28 Holstein calves from a dairy farm, located on an endemic region for anaplasmosis, by IB-ELISA and rMSP5 ELISA showed a decrease in the mean ODs from day 7 to day 30, when the lowest ODs were recorded. From day 30, there was an increase in the ODs that peaked in the 360th day. This pattern was similar for both ELISAs, however the mean OD was always above the cut-off level in the IB-ELISA, whereas it reached the cut-off values in the days 15 and 30 in the rMSP5 ELISA (Fig. 4). DISCUSSION The high degree of identity of MSP5 between the Brazilian and the American isolates of A. marginale at the DNA and protein levels is in accordance with the high level of conservation of this protein in Anaplasma spp. reported previously (Visser et al. 1992, Reyna-Bello et al. 1998). The 1.58% of non-identity (10/633 bp) was possibly due to mutations and/or to random errors during amplification with Taq DNA polimerase, which has a error rate of 0.2 to 2 x 10-4 misincorporations per bp per cycle (Bracho et al. 1998). The purification of A. marginale IB was partially achieved, as some contamination with erythrocyte proteins was detected in the SDS-PAGE. On the other hand, it was possible to purify rMSP5 by electroelution. The choice of this method was due to the failure in the purification of rMSP5 by affinity chromatography in agarose/nickel columns. The identity of rMSP5 was confirmed by Western blot with MoAb ANAF16C1 (Visser et al. 1992). This MoAb also recognized MSP5 in the IB preparation, along with ANA22B1, which recognized MSP1a, confirming the conservation of these proteins during the purification steps of the IB. The higher molecular mass of rMSP5 (22 kDa) when compared with the native protein was expected, as the vector pTrcHis-TOPO encodes a 3-4 kDa N-terminal peptide, which includes a histidine tag. In regards to the performances of rMSP5 and IB ELISAs, there were no statistical differences neither in the sensitivity nor in the specificities. Also, there was no statistically significant difference in the rates of cross-reactions with B. bovis antibodies between the two ELISAs. This indicates that contamination of IB antigen with erythrocyte proteins did not influence the performance of the IB ELISA. Crude antigen ELISAs with erythrocyte proteins has been incriminated for false-positive reactions, as result of reaction with anti-erythrocyte antibodies in the sera (Barry et al. 1986). However, the crossed reactions probably occurred due to host immune reaction, because previously ELISA for detection of A. marginale antibodies displayed false positive reactions during acute phase of Babesia spp. infection (Madruga et al. 2000).Therefore, these ELISAs could be used for serological surveys in endemic areas. The performances of IB and rMSP5 ELISAs in detecting antibodies against A. marginale in an extensively raised cattle herd of Mato Grosso do Sul, Brazil, were very similar and characterized the region as enzootically-stable. In such epidemiological conditions, the risk of outbreaks of anaplasmosis is low. However, the sampling of animals with varying ages may underestimate the true status, as there is the possibility that some animals were infected at ages above nine months, when they are less resistant to tick-borne diseases (Mahoney 1977). The performances of IB and rMSP5 ELISAs in detecting antibodies against A. marginale in calves naturally-infected with A. marginale in a dairy farm were very similar. In both tests, maternal antibodies reached the lowest levels by 15-30 days, with an increase in the mean ODs in both ELISAs thereafter, suggesting the development of active immunity against A.marginale. The ODs were higher in the IB ELISA, remaining above the cut-off level all the time, whereas in the rMSP5 ELISA, the ODs reached the cut-off level on days 15 and 30. Results showed that all calves were seropositive by one-year old, characterizing a situation of enzootic stability. Despite the fact that calves were kept in individual stalls from day 2 up to four months of age, they were infected, since the ELISA ODs increased constantly. They were probably infected by ticks during the first day of life, when they remained with their mothers, or by blood-sucking diptera. The similar performances of the ELISAs suggest that both tests can be used in epidemiological surveys for detection of antibodies to A. marginale in cattle. However, the antigen production and the standardization of each batch of antigen seems to be easier in the rMSP5 ELISA. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06086t3.jpg] [oc06086t2.jpg] [oc06086f2.jpg] [oc06086t1.jpg] [oc06086f1.jpg] [oc06086f3.jpg] [oc06086f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}