 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 101, No. 5, August, 2006, pp. 543-546 Dynamics of feeding and defecation in Triatoma vitticeps (Stal, 1859) (Hemiptera, Reduviidae, Triatominae) and its potential in the transmission of Trypanosoma cruzi Claudiney Biral dos Santos/+, Gustavo Rocha Leite, Paulo Augusto Sessa, Aloísio Falqueto* Núcleo de
Entomologia da Secretaria de Saúde do Estado do Espírito Santo
*Unidade de Medicina Tropical, Universidade Federal do Espírito Santo,
Av. Marechal Campos 1468, 29040-090 Vitória, ES, Brasil Received 2 February
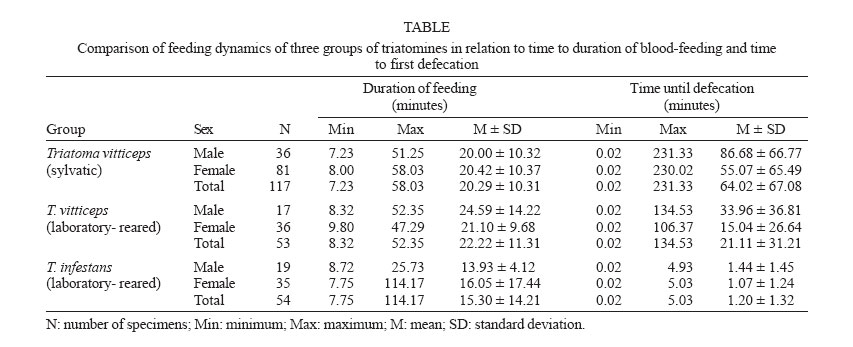
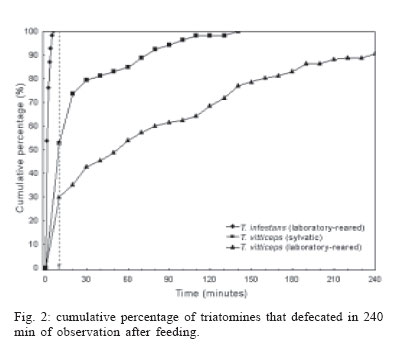
2006 Cod eNumber: oc06091 Adults of Triatoma vitticeps infected by flagellates similar to Trypanosoma cruzi are frequently captured by the inhabitants of rural areas in the Brazilian state of Espírito Santo. The dynamics of feeding and defecation were observed in three groups of adult triatomines, consisting of sylvatic T. vitticeps and laboratory-reared specimens of this species and T. infestans. Triatomines were observed from the moment they were presented with an immobilized chicken as a bloodmeal source until 240 min after feeding had ended. Mean times between the end of feeding and defecation for T. infestans, wild T. vitticeps and laboratory-reared specimens of the latter species were 1.2, 21.1, and 64 min respectively. All T. infestans defecated within 10 min of feeding, while only 29.9 of wild and 52.8% laboratory-reared specimens of T. vitticeps did so within this period. These results may explain the low efficiency of T. vitticeps in T. cruzi transmission to man. The shorter time between feeding and defecation in laboratory-reared T. vitticeps may indicate a change in behaviour of this species as a result of adaptation to an artificial environment. Key words: Triatoma vitticeps - feeding behaviour - defecation - vector potential - Trypanosoma cruzi The triatomine bug Triatoma vitticeps (Stal, 1859) occurs in the Brazilian states of Bahia, Espírito Santo, Minas Gerais, and Rio de Janeiro (Galvão et al. 2003). It is essentially a forest species, whose adults invade houses attracted by light or in search of blood meals, although they are apparently unable to colonize human habitations (Santos et al. 2005). In Espírito Santo, insects captured indoors present high rates of infection by flagellates morphologically similar to Trypanosoma cruzi (Chagas, 1911) (Sessa & Carias 1986, Dias et al. 1989, Santos et al. 2006). Among the characteristics that define the capacity of a triatomine to infect a host with T. cruzi are feeding and defecation behavior. Species that defecate during or shortly after taking a bloodmeal, so that infected faeces are deposited on the host, show the greatest vector potential (Dias 1956, Diotaiuti et al. 1995). It is known that T. vitticeps has low vector potential, due principally to the relatively long interval between feeding and defecation. The objective of the present study was to determine the dynamics of feeding and defecation in this species and compare it with Triatoma infestans (Klug, 1834), considered to be the most efficient vector of T. cruzi. MATERIALS AND METHODS Experiments were carried out in 2001-2003, using three groups of adult triatomines. The first consisted of 117 sylvatic specimens of T. vitticeps, the second 53 laboratory-reared individuals of this species and the third 54 specimens of T. infestans, also reared in the laboratory. Sylvatic specimens of T. vitticeps were captured by local inhabitants of houses in areas close to forest remnants in 81 localities of 27 municipalities of Espírito Santo and sent by the Regional Sanitary Units to the entomology laboratory of the State Health Service (Fig. 1). Insects were observed for two days and only those that appeared undamaged and active used in experiments. Triatomines of the second group were the first generation of laboratory-reared specimens coming from sylvatic triatomines collected in the state of Espírito Santo. The specimens of the third group were from colonies maintained in the laboratory for two generations coming from eggs supplied by the entomology laboratory of the Uni-versidade Federal de Goiás. The adults of the second and third groups were only used in the experiment seven days after having completed the ecdisis. Triatomines were held individually in mesh-covered pots containing a folded piece of filter paper. During a two years period, specimens of the three groups were exposed concomitantly to immobilized chickens used as alimentary source, until the complete bloodmeal. The insects were observed individually from the time they were exposed to the bloodmeal source until 240 min after the end of feeding. The following variables were controlled: duration of blood-feeding; interval between end of meal and first faecal deposition. ANOVA and chi-squared tests were used to compare the means and frequencies respectively, the level of significance being set at p ≤ 0.05. RESULTS Mean engorgement time of T. infestans was significantly shorter than that of the sylvatic (p = 0.026) and laboratory-reared (p = 0.006) T. vitticeps. The interval between feeding and the first faecal deposition were significantly different between the three groups, being shorter in T. infestans, with a mean of 1.2 min, compared to 21.1 and 64 min for laboratory-reared and sylvatic T. vitticeps, respectively (Table). All the T. infestans specimens defecated within 10 min of completing a bloodmeal, while only 52.8 of laboratory-reared and 29.9% of sylvatic T. vitticeps specimens did so within this interval (Fig. 2), and the differences between the three groups were highly significant (χ2 = 72.8; p < 0.001). Of the 117 T. vitticeps sylvatic specimens, 11 did not defecated in the observation time of 240 min. The mean times to defecation of females were shorter than those of males of their respective groups, but without statistic significance. The percentage of males that did not defecate within 10 min of completing a bloodmeal was significantly higher than females (χ2 = 9.2; p = 0.002). Two females and one male of sylvatic T. vitticeps, as well as one laboratory-reared female defecated during engorgement. DISCUSSION According to Zeledón et al. (1977), any species of triatomine in which defecation occurs within 10 min of taking a bloodmeal has a significant chance of being a vector of T. cruzi. Other factors such as adaptation to the domestic environment, indoor density, affinity with the host, and density-dependent timing of defaecation are also important in determine its vector potential (Piesman & Sherlock 1983, Kirk & Schofield 1987, Trumper & Gorla 1991, Braga & Lima 1999). Dias (1956) compared the dynamics of defecation in six South American species of triatomine, including nymphs of T. vitticeps, observing that this species delayed defecation for longer than the others. Gonçalves et al. (1988), studying the life cycle of this species, also observed a relatively long delay in defecation among specimens reared in the laboratory. In the present study, comparative analysis of the three groups revealed that T. infestans took markedly less time to defecate after feeding, all the specimens of this group doing so within 10 min. In contrast, a low proportion of individuals of T. vitticeps defecated within 10 min, and the mean interval between feeding and defecation time was very high, factors which reduce the possibility of T. cruzi transmission. The differences observed between males and females, with respect to the time taken to defecate after feeding and the percentage of insects that defecated with 10 min after a bloodmeal may be explained by the fact that females generally ingest greater volumes of blood to guarantee in response to the demands of oviposition (Gon-çalves et al. 2000). When the amount of blood ingested is large, triatomines eliminate their faeces more rapidly due to the high water content of blood (Friend & Smith 1985). Nattero et al. (2002) also observed a correlation between the volume of blood ingested and the time to defecation, confirming that females are potentially more efficient in infecting vertebrates with T. cruzi. The T. infestans group presented a mean engorgement time that was significantly shorter, probably due to its voracity and smaller size, resulting in shorter exposure to possible aggressive reactions by the host. Such adaptations guaranteed the success of this species in colonizing artificial ecotopes (Diotaiuti et al. 1995). Triatomines in the laboratory-reared group of T. vitticeps defecated sooner after feeding than wild-caught specimens, indicating an important behavioral modification. Any such change that was associated with adaptation to an artificial ecotope would be epidemiologically significant, increasing the vectorial capacity of the insect. According to Silveira et al. (1983), T. vitticeps adapts poorly to artificial ecotopes despite its high dispersal capability. Adult insects from sylvatic environments are captured quite frequently in houses, but colonization by nymphs is rare. The results of the present study, together with the inability of the insect to colonize houses, may explain the low indices of human infection with T. cruzi in rural populations of Espírito Santo. On the other hand, bloodmeal analysis by the precipitin method demonstrated the marked presence of human blood in adult insects captured in houses, indicating frequent contact of T. vitticeps with man (Dias et al. 1989, Gonçalves et al. 2000). This, together with the high rate of natural infection by flagellates similar to T. cruzi in adult insects captured in houses, highlights the need for epidemiological vigilance to reduce the risk of T. cruzi transmission to man. ACKNOWLEDGEMENTS To Arlinete Souza de Medeiros and Waldomir Alves Bezerra for providing T. infestans eggs to set up the colony and to Bruce Alexander for translating the manuscript to English. REFERENCES
The following images related to this document are available:Photo images[oc06091f2.jpg] [oc06091t1.jpg] [oc06091f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}