 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 101, No. 5, August, 2006, pp. 547-550 Susceptibility of Anopheles aquasalis and An. darlingi to Plasmodium vivax VK210 and VK247 Ana NM da Silva, Carla CB Santos*, Raimundo N Lacerda*, Ricardo LD Machado**, Marinete M Póvoa*/+ Núcleo de
Medicina Tropical, Universidade Federal do Pará, Belém, PA,
Brasil *Laboratório de Pesquisas em Malária, Seção
de Parasitologia, Instituto Evandro Chagas, Br 316, km 7 s/no,
67030-000 Ananindeua, PA, Brasil **Departamento de Doenças Infecciosas
e Parasitárias, Faculdade de Medicina, São José do Rio
Preto, SP, Brasil Financial support: Evandro Chagas Institute project "Comparative Susceptibility of Anopheles aquasalis and An. darlingi to Plasmodium vivax VK210 and VK247" Received 2 February
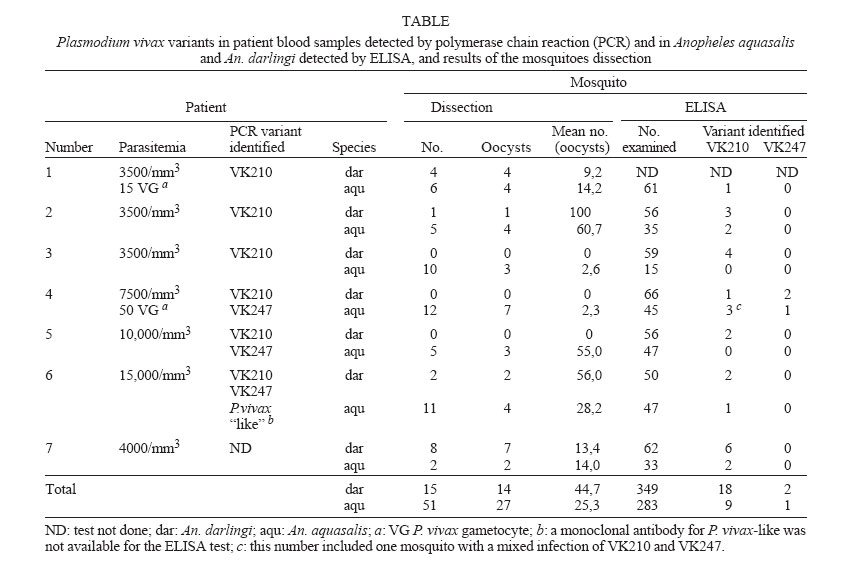
2006 FF Code Number: oc06092 The susceptibility of Anopheles aquasalis (F3 generation) and An. darlingi (F1 generation) to Plasmodium vivax circumsporozoite protein phenotypes from a limited number of blood samples of malaria patients in Belém, state of Pará, Brazil, was examined. A polymerase chain reaction was used to determine the P. vivax phenotypes in blood samples and the blood-fed infected mosquitoes were dissected and tested by ELISA. In all patient infections, more infected An. aquasalis and An. darlingi were positive for VK210 compared with VK247. Key words: malaria - Plasmodium vivax - VK210 - VK247 - Anopheles aquasalis - Anopheles darlingi - Brazilian Amazon In Belém (the capital of the state of Pará) two species have been incriminated as important malaria vectors (Davis 1931, Galvão et al. 1942, Póvoa et al. 2003): Anopheles (Nyssorhynchus) aquasalis Curry 1932, a coastal Neotropical species (Zimmerman 1992), considered to be the primary coastal malaria vector of Plasmodium vivax in Venezuela (Berti et al. 1993) and as far as Southeastern Brazil (Forattini 1962); and An. darlingi Root 1926, a Neotropical species that is highly susceptible to Plasmodium and the primary vector in the Amazon (Deane et al. 1948, Klein et al. 1991). In Brazil overall, most malaria cases are caused by P. vivax (Akhavan et al. 1999), but the proportion of P. vivax to P. falciparum varies regionally (Brazilian Ministry of Health 2000). In Pará, from 1985 to 1999 the number of malaria infections by P. vivax and P. falciparum differed significantly (p = 0.003) with the incidence of P. vivax being considerably higher (Póvoa et al. 2003). From 1993-1999, in Belém, the proportion of malaria cases identified by Giemsa-stained blood smears as P. vivax fluctuated from a low of 85% in 1994 to a high of 97% in 1997 and 1998 (Póvoa et al. 2003). In all likelihood, in Belém, these parasites were transmitted by An. aquasalis and An. dar-lingi (Póvoa et al. 2003). Based on the repeat units of the circumsporozoite (CS) proteins, three P. vivax variants have been identified: VK210 (amino acid sequence is GDRA(D/A)GQPA) (Arnot et al. 1985), VK247(amino acid sequence is ANGA(G/D)(N/D)QPG) (Rosenberg et al. 1989), and P. vivax-like (amino acid sequence is APGANQ(E/G)GGAA) (Qari et al. 1993). All three variants are present in the Brazilian Amazonian region, including Belém (Arruda et al. 1996, 1998, Machado & Póvoa 2000). However, it is rare to detect VK247 as a single infection; this variant is generally found in mixed infections with either VK210 or with both VK210 and P. vivax-like (Machado & Póvoa 2000, Machado et al. 2003). Recent studies of Neotropical anophelines have described differential susceptibilities of An. albimanus and An. pseudopunctipennis to P. vivax variants VK210 and VK247 (González-Ceron et al. 1999, Rodriguez et al. 2000), but there have been no previous studies of either An. aquasalis or An. darlingi susceptibilities to P. vivax variants. Klein et al. (1991) found that An. darlingi from Ron-dônia in western Amazonian Brazil (among several other anopheline species) was very susceptible to P. vivax infection. More recently, F1 An. konderi, An. oswaldoi, and An. darlingi collected in Acre and the state of Rondônia, Brazil, were all shown to be susceptible to infection by P. vivax (Marrelli et al. 1999). However, in both studies the authors did not know which P. vivax circumsporozoite protein phenotypes the mosquitoes speciemes were dealing with. Our objective was to examine the susceptibility of An. aquasalis and An. darlingi from Belém to P. vivax VK210 and VK247. MATERIALS AND METHODS Patients - Seven subjects diagnosed with P. vivax malaria by Giemsa-stained blood smears at Instituto Evandro Chagas (IEC) or at Pará health clinics, were the source for mosquito infection. All individuals were older than ≥ 21 years old, infected with circulating asexual stages of P. vivax, parasitemia higher than 3000 parasites/mm3 and with history of previous malaria or not. The exclusion criteria were: individuals with debilitating symptoms such as vomit, diarrhea, dehydration caused by the infection, pregnant women, children, and indigenous people. The history of previous malaria episodes, approximate date of initial symptoms, and probable site where malaria transmission occurred were obtained from each patient. Treatment with chloroquine and primaquine, alone or combined was provided to each patient as soon as the mosquito feeding was completed. This study was approved by the Ethical Committee on Human Subjects at the IEC, protocol number CEP/IEC 06/99. Infected mosquitoes - A maximum of 70 females, 3-6 days old laboratory reared An. aquasalis -F3 (Silva et al. 2006) and An. darlingi (F1), both obtained from females collected in Belém, Pará, were fed simultaneously for up to 30 min on the arms and legs of a same patient between to 2-11 days after the first report of symptoms. Mosquitoes were not fed during 12 h prior the infected blood feeding. After complete engorgement, the mosquitoes were maintained at 23-28oC and 70-80% RH, and provided with 10% sucrose ad libitum. Mosquito dissections were conducted on days 8-10 post blood meal. After staining the midguts with 1% mercurochrome (Eyles 1950), oocysts were counted at 400x using a light microscope. Identification of Plasmodium vivax variants in patients - Identification of the variants was done using GFM-PCR-ELISA as described in Machado and Póvoa (2000). As a source of DNA, 1/8 of a drop of blood (Warhurst et al. 1991) was placed in 5 µl of 10 x 1M buffer (tris pH 8.3, 0.01% w/v gelatin, 10 M potassium chloride, and 0.0015 M magnesium chloride); 2.5 µl of each oligonucleotide [AL60 (GTC GGA ATT CAT GAA GAA CTT CAT TCT C) and AL61 (CAG CGG ATC CTT AAT TGA ATA ATG CTA GG)] (Qari et al. 1993); 1 µl of each dNTPs (2.5 µmol Pharmacia Biotech) to a final concentration of 200 µM; 0.25 µl of Taq (Biotaq 5U/µl units Bioline M95601B) and 30.75 µl of sterilized distilled water. As a negative control we used two samples of uninfected blood in each time we did a GFM-PCR-ELISA identification. Detection of infection in mosquitoes - Ten to twelve days post infected blood-fed mosquitoes were killed and the Plasmodium variant determined by ELISA following the protocols in Wirtz et al. (1991, 1992). Heads/thoraces of An. darlingi and An. aquasalis that blood fed in each patient were tested. Negative controls consisted of males of each species. Positive controls were those provided by Kiekegaard & Perry Laboratories, US. The cut-off was two times the mean values of the optical density of the negative controls. Statistical analysis - The differences in susceptibility to infection between the two anopheline species and the two P. vivax variants were tested by Mann-Whitney. The significance level for statistical inference was p < 0.05. RESULTS The parasitemia for patients 1 and 4 -7 was detected at the IEC clinic and patient 2 and 3 at the Health Care Clinics and confirmed at the IEC, and ranged between 3500-15,000 parasites/mm3. Only in patient 1 and 4 were identified gametocytes (Table). In this study, by chance, six out of seven patients in the study all became infected in different parts of Pará (Anajás, Concórdia do Pará, Igarapé-Miri, São Caetano de Odivelas, São Luiz de Igarapé Açu, and Vizeu). Patients 1 and 4-6 had no previous history of malaria. Patient 2 have had malaria twice; patient 3, five times; and patient 7, four times previously. The variants present in each patient, determined by polymerase chain reaction, included VK210 (3 patients), VK210/VK247 (2 patients), and VK210/VK247/P.vivax-like (1 patient; Table). The P. vivax variant was not determined for patient 7, but the ELISA results of the infected An. aquasalis and An. darlingi show that this person had at least VK210 (Table). Comparing all the mosquitoes positives by ELISA, we found significant difference between the two mosquitoes species (p < 0.001) [mean number of mosquitoes: An. darlingi (3.33) and An. aquasalis (1.28)], as well as between the number of both An. aquasalis and An. darlingi infected with VK210 versus VK247 (p = 0.0012) (Table). Of the mosquitoes that only fed upon patients with mixed infections (patients 4-6; Table), all were susceptible to infection with P. vivax VK210, except An. aquasalis on patient 5, who had a parasitemia of 10,000 parasites/ mm3. There was a higher proportion of An. aquasalis and An. darlingi infected with VK210 (2.89) versus VK247 (0.96) in mixed infections (patients 4 -6; Table). However, in patient 4, who had a parasitemia of 7500 parasites/mm3 and 50 gametocyte forms, both An. aquasalis and An. darlingi became infected with VK210 and VK247 (Table). We were unable to evaluate mosquito susceptibility to infection with P. vivax VK247 because these variants were never found alone, only in mixed infections in Belém (Machado & Póvoa 2000). The dissection of the midgut of An. darlingi and An. aquasalis, demonstrated that both species developed infection by P. vivax, but the proportion of infected An. darlingi (93.33%) and the mean number of oocysts (44,65 ± 42,52) was higher than An. aquasalis (52.94%) mean number (25,28 ± 23,94; Table). On some dissected mosquitoes of both species, infected by mixed infection carriers, we had observed degenerated oocysts (data not showed). DISCUSSION Our results demonstrate for the first time, experimentally, that An. aquasalis from Belém are susceptible to P. vivax VK210 and VK247 and confirm the susceptibility of An. darlingi to P. vivax variants (Klein et al. 1991, Marrelli et al. 1999). We have previously demonstrated that the percentage of An. aquasalis naturally infected by P. vivax has increased in Belém from 0.26% (Galvão et al. 1942) to 1.18% (Póvoa et al. 2003). Our results suggest that additional regional comparative studies on Anopheles susceptibility to Plasmodium infections that include An. darlingi and the identification of P. vivax variants in both the patients and the infected mosquitoes would contribute significantly toward a better understanding of malaria epidemiology and parasite transmission rates in Brazil. In mixed infections both An. aquasalis and An. dar-lingi appear to become more infected with VK210 rather than to VK247, but in one patient infected with VK210/VK247 (patient 4) we detected VK210 and VK247 in both mosquito species. Our results may have been influenced by the small sample size. However, several studies have shown differences on the infectivity of anofelines to the variants. In Mexico, An. albimanus is more susceptible to infection with VK210, and An. pseudopunctipennis is more susceptible to VK247 (González-Ceron et al. 1999, 2001). Furthermore, in Southern Mexico, the prevalence of these two variants is closely associated with the distribution of these two mosquito species (Rodriguez et al. 2000). Our study is suggestive that a similar phenomenon, observed for An. albimanus (greater susceptibility to VK210) may occur in both An. aquasalis and An. darlingi, in Amazonian Brazil. We hypothesize that the greater probability of VK210 sporozoites development in An. aquasalis and An. darlingi compared with VK247 is either determined by the presence of a greater proportion of VK210 gametocytes circulating in patients or by other mechanisms (Vlachou & Kafatos 2005) such as ookinete destruction and (or) oocyst development arrest as observed for An. albimanus that is resistant to infection with VK247 (González-Ceron et al. 2001). These mechanisms may explain in the preferential development of VK210 in An. aquasalis and An. darlingi. Studies on mosquitoes experimentally infected in blood donors from regions of Colombia demonstrated that despite of the higher prevalence of anti-VK210 on these individuals, the mosquitoes produced more abundantly sporozoites VK247, suggesting that the anti-VK210 antibodies was blocking the development of sporozoites VK 210 (González et al. 2001). In contrast, our study do not allow us to conclude about the action of specific antibodies (anti- VK210 or VK 247) on the mosquitoes infection or the intensity of this infection, first because of our sample size and second, three (4-6) of our seven donors had no previous malaria. Further investigations are important to assess the variants immunogenicity as well as the infection in mosquitoes. Susceptibility is a condition in which the body tissues of the insect can be successfully infected by the parasite (Sinden 2002), thus our findings indicate that An. darlingi and An. aquasalis were susceptible to variants P. vivax. However, we can not compare the intensity of the infection, since the number of mosquitoes analyzed was variable. In general, An. darlingi was proportionally more infected than An. aquasalis. Obviously, the mechanisms of interaction parasite/mosquito are different among the species (Sinden 2002), but we had observed difference in feeding time and blood engorgement (An. darlingi was more efficient than An. aquasalis) which may have further implications in their vector competence (Chadee & Beier 1995, Takken et al. 1998). ACKNOWLEDGEMENTS To RA Wirtz for providing anti-P.vivax PK 210 and PK 247 monoclonal antibodies; and Jan E Conn for editorial and scientific comments on this manuscript. REFERENCES
The following images related to this document are available:Photo images[oc06092t1.jpg] |
| |||||||||

{kind=link}