 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101, No. 6, September ,2006, pp. 597-602 Detection of rifampin resistance patterns in Mycobacterium tuberculosis strains isolated in Iran by polymerase chain reaction-single-strand conformation polymorphism and direct sequencing methods Bahram Nasr Isfahani/+, Akbar Tavakoli, Mansoor Salehi*, Mehdi Tazhibi** Medical Microbiology Department *Molecular Biology
Department, School of Medicine **School of Health, Isfahan University
of Medical Sciences, Isfahan, Iran Financial support: Isfahan University of Medical Sciences Received 16 January 2006 Code Number: oc06102 Mutations in the rpoB locus confer conformational changes leading to defective binding of rifampin (RIF) to rpoB and consequently resistance in Mycobacterium tuberculosis. Polymerase chain reaction-single-strand conformation polymorphism (PCR-SSCP) was established as a rapid screening test for the detection of mutations in the rpoB gene, and direct sequencing has been unambiguously applied to characterize mutations. A total of 37 of Iranian isolates of M. tuberculosis, 16 sensitive and 21 resistant to RIF, were used in this study. A 193-bp region of the rpoB gene was amplified and PCR-SSCP patterns were determined by electrophoresis in 10% acrylamide gel and silver staining. Also, 21 samples of 193-bp rpoB amplicons with different PCR-SSCP patterns from RIFr and 10 from RIFs were sequenced. Seven distinguishable PCR-SSCP patterns were recognized in the 21 Iranian RIFr strains, while 15 out of 16 RIFs isolates demonstrated PCR-SSCP banding patterns similar to that of sensitive standard strain H37Rv. However one of the sensitive isolates demonstrated a different pattern. There were seen six different mutations in the amplified region of rpoB gene: codon 516(GAC/GTC), 523(GGG/GGT), 526(CAC/TAC), 531(TCG/TTG), 511(CTG/TTG), and 512(AGC/TCG). This study demonstrated the high specificity (93.8%) and sensitivity (95.2%) of PCR-SSCP method for detection of mutation in rpoB gene; 85.7% of RIFr strains showed a single mutation and 14.3% had no mutations. Three strains showed mutations caused polymorphism. Our data support the common notion that rifampin resistance genotypes are generally present mutations in codons 531 and 526, most frequently found in M. tuberculosis populations regardless of geographic origin. Key words: Mycobacterium tuberculosis
- rpoB gene mutation - polymerase chain reaction-single-strand
conformation
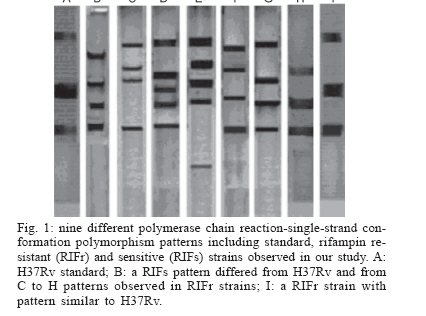
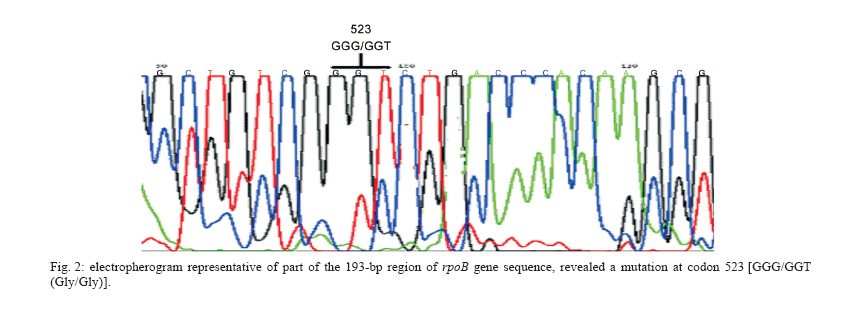
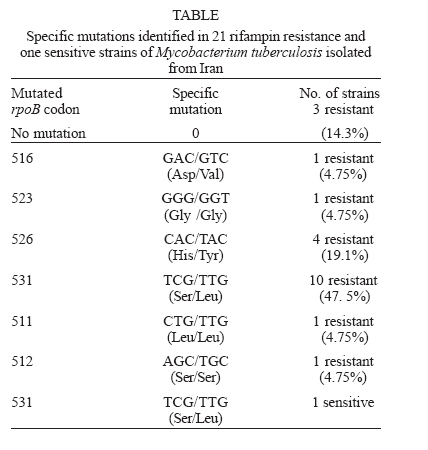
Mycobacteria are responsible for considerable human morbidity and mortality worldwide (Bahremand 1992). In the last decade, tuberculosis (TB) has reemerged as one of the leading causes of death with nearly 3 million deaths annually (Bloom & Murray 1992). The emergence of AIDS and decline of socio-economic standards contribute to the disease's resurgence in industrialized countries (Frieden et al. 1993, Ashok et al. 1998). A more alarming aspect of tuberculosis is the emergence of multidrug-resistant strains of M. tuberculosis (MDRTB) that is a widespread phenomenon (Snider & Roper 1992, Frieden et al. 1993, Iseman 1993, Kato-Maeda et al. 1999). The common finding has been a high rate of primary resistance to isoniazid (INH) and to the combination of isoniazid and rifampin (RIF) (Garcia-Garcia 2000) and a high rate of resistant (17.2%) to all the four commonly used antimycobacterial agents for treatment and prophylaxis of TB, including RIF (Granich et al. 2000, Singh & Kaur 2004). Prevalence rate of TB in 5 years ago in Iran were reported as high as 141 in 100,000 in Zabol (Khalil 2003) and as low as 11.9 in 100,000 in Isfahan (Zandeeh et al. 2003). Because of its high bactericidal action, RIF, along with INH, forms the backbone of short-course chemo-therapy. Resistance to RIF is increasing because of widespread application and results in selection of mutants resistant to other components of short-course chemotherapy; 88% of hospitalized patients with drug resistant TB in Massih Daneshvari Hospital (Iran-Tehran) were resistant to at least INH and RIF (Masjedi et al. 2003). In 56% the TB bacillus was resistant to all four drugs: INH, RIF, streptomycin (STM), and ethambutol (EMB); 85% had resistance to STM at least and 60% were at least resistant to EMB. Most drug resistant cases of TB in were seen among Afghan emigrants; 95% of cases had a history of treatment at least once, and the resistance was secondary; 36% of MTB strains isolated from patients in Mshhad-Iran were resistant to at least one anti-TB drug (Namaei et al. 2003). The highest rate of resistance was observed for SM: 34.29 and 60% of strains isolated from newly and previously treated patients respectively were resistant to streptomycin; 4% were multidrug resistant (MDR). Characterization of the rpoB gene (a region responsible to synthesis of beta-subunit of RNA polymerase) in Escherichia coli demonstrated that RIF specifically interacts with this subunit; thereby hindering transcription, and mutations in the rpoB locus conferred conformational changes leading to defective binding of the drug and consequently resistance (Jin & Gross 1988). Several techniques were used for detecting these mutations and subsequently, the rpoB locus from M. tuberculosis was characterized and mutations conferring the resistant trait were identified (Telenti et al. 1993a, Kapur et al. 1994). Automated sequencing has been unambiguously applied to characterize mutations associated with resistance to RIF, but a number of other techniques such as PCR-single-strand conformational polymorphism (PCR-SSCP), have been successfully applied (Pretorius et al. 1995, Morris et al. 1995, Kalia et al. 1997). PCR-SSCP analysis, involves amplification of a segment of the gene encoding for the specific drug target and comparison of PCR products of drug-sensitive and drug-resistant strains by non-denaturing electrophoresis, in which mutations usually result in an altered pattern (Snider & Roper 1992, Iseman 1993, Telenti et al. 1993b). However recent studies have questioned its sensitivity and specificity (Lee et al. 1998, Miriam et al. 2001). Also, from various region of the world, different types of mutations were identified in RIFr M. tuberculosis clinical isolates. Most of these were point mutations involving 10 codons, while only few isolates had a deletion or insertion. Most frequently involved in mutation in the various regions of the world were codon 531 and codon 526 (Matsiota et al. 1998, Lee et al. 1998, Spindola et al. 2001, Miriam et al. 2001). In this study, we investigated the usefulness of PCR-SSCP method to detect mutations in the rpoB gene of RIFr and RIFs M. tuberculosis strains isolated in Iran from 2002 to 2004. Also the frequencies of mutations in their rpoB gene were characterized by direct sequencing to determine the codons most frequently involved in resistance to RIF in Iran. MATERIALS AND METHODS In the present study, 37 M. tuberculosis strains, originating from the Iranian Tuberculosis Center were used; they had been sent from laboratories and TB centers in Iran from 2002 to 2004. All isolates were characterized by conventional methods (Staining by Ziehl Neelsen method, observation of speed of growth and colony morphology and nitrate reduction test) and by specific amplification of the Direct Repeat (DR) gene region only present M. tuberculosis complex (Bahremand 1992, Tavakoli et al. 2005). The indirected agar proportion method test used for testing the resistance or sensitivity of pure cultures of M. tuberculosis isolates (Williams et al. 1998, Charley et al. 1999). All isolates were cultured in 7H9 broth medium for ten days. Subsequently, LJ medium without and with RIF were inoculated with 0.1 ml of 10-2 and 10-4 dilutions of a McFarland 1.0 concentration of a suspension of each isolate of M. tuberculosis. The inoculated plates were then incubated at 37°C for three weeks. The break-point was 1%. The isolate was considered susceptible to RIF if the number of colonies that grew on the drug-containing plates were more than 1% of the number of colonies that grew on the RIF-free control. An isolate was considered resistant if 1% or more grew on the drug-containing plate. Chromosomal DNA of 37 clinical isolates, 21 RIFr and 16 RIFs, were extracted by "DNA mini-prep" procedure using Hexadecyltrimethylam-moniumbromide (CTAB) as previously described (Cooper et al. 1999, Spindola et al. 2001). About 10 ng of extracted DNA was used for PCR amplification. The DNA of drug-susceptible standard strain, H37Rv (ATCC 27294) was used as control sample in this study. PCR was performed as following: PCR reactions (50 µl) contained 10 ng target DNA, 15 pmol primers, 2 mM dNTP (Pharmacia Biotech), 2.5U Taq polymerase (MBI Fermentas), 1.5 mM MgCl2, and 5 µl 10 × buffer. The reaction was performed in a thermal cycler from PCR Hybid (Omnigene). The sequence of primers for the rpoB locus were (rpo 105) 5'-CGTGGAGGCGATCACACCGCAGACGT-3' and primer (rpo 273) 5'-gacctccagcccggcacgctcacg-3', which produced a 193-bp amplicons (Spindola et al. 2001). Samples were then subjected to one cycle of 96°C for 5 min, followed by 40 cycles of 96°C for 1 min, 55°C for 1 min, 72°C for 30 s, and one final cycle of 72°C for 10 min. PCR products were run on 2% agarose gel and examined for the presence of the 193-bp band after ethidium bromide staining (Travis 1995, Miriam et al. 2001). The PCR products were SSCP analyzed by electrophoresis on 10% (w/v) acrylamide gels. In brief, the SSCP gel was made by mixing 10 ml 40% (w/v) acrylamide solution, 25.6 ml H2O, 4 ml 10 × TBE, 30 µl TEMED, and 300 µl ammonium persulfate. About 6 µl of amplified product was mixed with 4 µl of loading buffer (95% formamide, 20 mM EDTA, and 0.05% bromophenol blue) and the mixtures was boiled for 4 min, cooled on ice for 5 min, and then loaded on the acrylamide gel at 2 W for 14-16 h in cold room. The gel was silver stained by using 10% (v/v) gacial acetic acid as fix/stop solution; silver nitrate solution (contained 2 g AgNO3, 2 l ultrapure water and 3 ml 37% formaldyde) as stain solution and solution contained 60 g sodium carbonate, 2 l ultrapure water, 3 ml 37% formaldyde and 400 µl sodium thiosulfate as developing solution. SSCP assays were repeated at least two times with all isolates. Twenty one samples of 193-bp rpoB amplicons with different PCR-SSCP patterns from RIFr (resistance) and 10 RIFs (sensitive) were sequenced. The products were sequenced by using upstream primer rpo105 with Applied Biosystems 377 automated sequence protocol (ABI PRISMTM Dye Terminator). All post runs analysis was performed using Clustalw, version 1.82, and software, EMBL-EBI. Each sequence was compared both with the control strain sequence and with the published rpoB sequence (GenBank accession NC 000962 REGION: 759805.763323). RESULTS The results of susceptibility testing indicated that percentage resistance were from less than 1 to 31 on LJ medium contained 4 µg RIF ml-1. This percentage resistance were less than 1 for all sensitive isolates. Preparations of inoculums were problematic, because of over- or under inoculation, so that we had to repeat the experiment for more than six times in about 8 months. The PCR-SSCP patterns from strains is shown in Fig.1. On the basis of the SSCP results, the 21 RIFr isolates were grouped in seven groups: pattern 1, 4, and 5 with 4 bands; pattern 2 and 3 with 5 bands; pattern 6, similar to H37Rv; and pattern 7 with 3 bands. Of the16 RIFs, 15 strains (93.8%) had a PCR-SSCP pattern identical to H37Rv; and 1 strain (6.2%) had different pattern. One of the resistant isolates had a PCR-SSCP pattern similar to that of M. tuberculosis susceptible control strain H37Rv and one of the RIFs strains had a PCR-SSCP pattern differ to that of susceptible control strain H37Rv. After SSCP analysis, 21 PCR product samples from RIFr and 10 from RIFs were sequenced. An electropherograms of automatic DNA sequencing is shown in Fig.2. Of the 21 sequences of RIFr isolates, 18 strains showed a single mutation in the 81-bp region of the rpoB gene. Three strains with 1 to 5-percentage resistance in 4 µg RIF ml-1 did not show any mutation within the 81-bp core region. 14.3% had mutations caused polymorphism and therefore should have another mutation not in the 81-bp region that needs to be characterized. Ten samples had a mutation at codon 531(Ser/Leu), four at codon 526 (His/Tyr), one at the codon 516 (Asp/Val). Three stains had polymorphism in codons, 523 (Gly/Gly), 512 (Ser/Ser), and 511 (Leu/Leu) with uncharacterized mutation in another region outside the 81-bp region and finally three strains did not have neither mutations nor polymorphism. These data are summarised in Table. Most of resistant strains (95.2%) showed PCR-SSCP patterns different from that of H37Rv; 28.6% had four bands and 23.8% had three bands. These finding demonstrated that frequencies of particular PCR-SSCP patterns in RMP-resistant M. tuber-culosis isolates from Iran have some differences from those that have been reported from other geographic areas (Kochi et al. 1993, Telenti et al. 1993a,b, Kim et al. 1997, 1999, Lee et al. 1998, Miriam et al. 2001). We repeated our experiments several times and the results demonstrated that our technique is highly reproducible; although it should be considered that differences in methods might produce different banding pattern. Nonetheless local differences, our data also demonstrated that SSCP analysis is highly specific and sensitive for detecting mutations in the rpoB gene in RIFr M. tuberculosis. The sensitivity of this method was 95.2% with the specificity of 93.8%. Thus this method can be considered as a convenient method for detection of mutations responsible for conferring RIFr. Our results are the same as to those of Bahremand et al. (1998) finding but are not exactly conform to Spindola de Miranda et al. (2001), demonstrating that this method may perform poorly in detecting mutations in rpoB region. However, in this study, 1/21 (4.75 %) of the resistant isolates had a PCR-SSCP pattern similar to that of M. tuberculosis susceptible control strain H37Rv. These results may imply a mutation in another part of the rpoB gene or the existence of at least one additional gene that participates in RIFr. Sequencing result of this strain demonstrated no mutation in the 81-bp region. Also 1/16 (6.3 %) of the sensitive strains had a PCR-SSCP pattern different from that of susceptible control strain H37Rv that may imply the validity of proportion method is lower than 100%. Surprisingly sequencing of this strain demonstrated codon 531 (TCG/TTG) mutation that cause (Ser/Leu) change. A study on RIFr detection of M. tuberculosis from Brazilian patients using reverse-phase hybridization assay was performed by de Oliveira et al. (2003). They found that although all sensitive strains had the sensitive genotype, but among resistant isolates, a sensitive genotype was obtained in three due to the absence of mutations in the hot spot region, demonstrating an assay accuracy of 97.6% for detection of drug susceptibility. They also reported that, using specific probe for the mutation; the sensitivity to detect heteroresistance is similar to that of the proportion method. Our PCR-SSCP analysis results are not fully consistent with that of reports from other geographic areas (Telenti et al. 1993b, Xiao-Yong et al. 2003, Tracevskr et al. 2002). On the basis of the SSCP results of Miriam et al. (2001), the 35 RIFr isolates were grouped in two main categories: group one, 24 isolates (68.6%) with an SSCP identical to that of the control strain H37Rv, and group two, 11 isolates (31.4%) with an SSCP different from that of H37Rv. In their study there also were 11 RIFs isolates that showed an SSCP identical to that of H37Rv. Lee et al. (1998) assessed the molecular mechanism of RIFr in clinical strains of M. tuberculosis. The molecular nature of a part of the rpoB gene in 77 M. tuberculosis clinical strains isolated in Korea was analyzed using PCR-SSCP and PCR-sequence analysis. Among 67 RIFr isolates, 50 showed SSCP profiles different from that of a RIFr control strain, M. tuberculosis H37Rv, while 17 resistant isolates displayed SSCP profiles indistinguishable from that of the sensitive control strain. This might be due to the presence of mutation outside of the amplicon. After sequencing of 81-bp region of the rpoB gene of 21 RIFr isolates, 85.7% strains showed a single mutation. An additional interesting finding was that three strains (14.3%) with 1 to 5-percentage resistance in a MIC of 4 µg RIF ml-1 did not show a mutation within the 81-bp core region. This observation confirms a recent report of a RIFr strain identified in Australia with a mutation outside the core region of rpoB (Lilly et al. 1999); 14.3% had mutations causing SSCP polymorphism, with a probable mutation in another region. Upon analysis of mutation frequency distribution, 47.6% had a mutation at codon 531 (Ser/Leu). In addition, 19% isolates had mutations at codon 526 (His/Tyr) and 4.7% at the codon 516 (Asp/Val) (Table). These finding is comparable to the results of early studies demonstrating the rpoB mutation frequencies in isolates from the United States (Kapur et al. 1994, Telenti et al. 1993a,b), European and African countries (Lilly et al. 1999, Hirano et al. 1999), Japan (Ohno et al. 1996, Suzuki et al. 1995), Brazil and France (Spindola et al. 2001) and Asian countries (Hirano et al. 1999). In summary, our data on rpoB mutation frequencies in isolates from Iran supported the common notion that rifampin resistance genotypes with mutations at critical codons, i.e., those encoding Ser-531 and His526 were the most frequently found in M. tuberculosis populations regardless of geographic origin. Mutations associated with nucleotide replacements in codons 513 and 526 were associated with high-percentage RIFr, whereas mutations in codon 523 and 512 were observed in low-percentage RIFr. Other authors (Kapur et al. 1994) have reported high and low levels of resistance associated with specific nucleotide replacements. These differences reflect the complex and crucial interaction between the drug and its target at the molecular level, where the position of the affected allele seems to be critical. ACKNOWLEDGEMENT To Dr Ghazisaeedi from Tehran University and Dr Karimi from Pasteur institute of Iran for scientific associations and providing mycobacterial strains. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06102f2.jpg] [oc06102t1.jpg] [oc06102f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}