 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101, No. 6, September ,2006, pp. 639-643 Primary culture of the region of the amebocyte-producing organ of the snail Biomphalaria glabrata, the intermediate host of Schistosoma mansoni L Barbosa, LM Silva*, PMZ Coelho**/***, SR Santos*/**, CL Fortes-Dias*/+ Departmento de Parasitologia, Universidade Federal
de Minas Gerais, Belo Horizonte, MG, Brasil *Fundação
Ezequiel Dias, Diretoria de Pesquisa e Desenvolvimento, Laboratório
de Biologia Molecular e Celular, R. Conde Pereira Carneiro 80, 30510-010
Belo Horizonte, MG, Brasil **Centro de Pesquisas René Rachou-Fiocruz,
Belo Horizonte, MG, Brasil ***Santa Casa de Misericórdia,
Belo Horizonte, MG, Brasil Financial support: Fundação Ezequiel Dias, Centro de Pesquisas René-Rachou, CNPq (with research fellowships to CL Fortes-Dias and PMZ Coelho) Received 8 March 2006 Code Number:oc06108 Biomphalaria glabrata snails are major hosts for the digenetic trematoda Schistosoma mansoni, the causative agent of human schistosomiasis. The success or failure of the infection will be dependent on the mobilization of the molluskan internal defense system, where a major role will be played by circulating hemocytes produced by the APO (amebocyte-producing organ) of the snail. In this report, the primary culture of the APO region of B. glabrata was obtained for the first time, as well as a control culture of the ovotestis. Three different cell populations migrated easily from the explants in culture, with no need of any dispersion agent. The cells grew in suspension at an incubation temperature of 15ºC and the cultures were maintained viable for up to two weeks. Two of these cell populations obtained resembled cell types known to be present in the hemolymph of Biomphalaria. The availability of APO cells in culture may contribute to a better understanding of the internal defense in mollusks, in general, as well as the specific response of B. glabrata to S. mansoni infection. Key words: Biomphalaria - schistosomiasis - Schistosoma mansoni - hemocyte - tissue culture
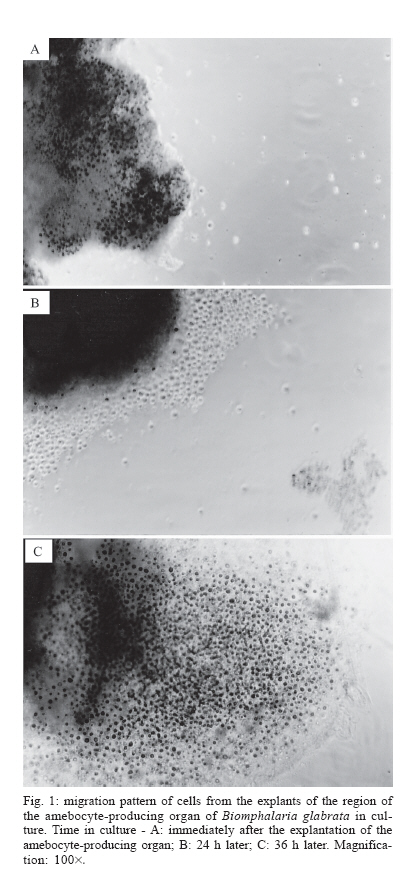
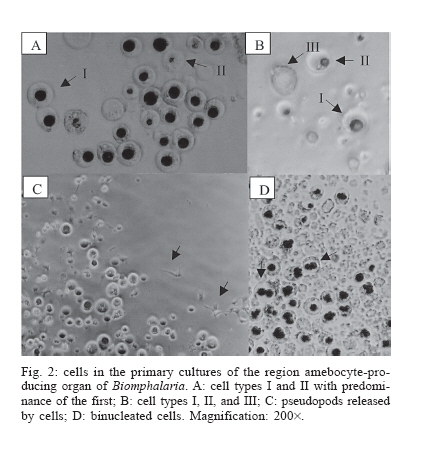
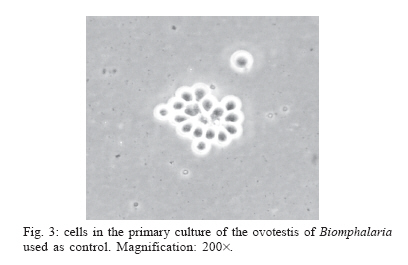
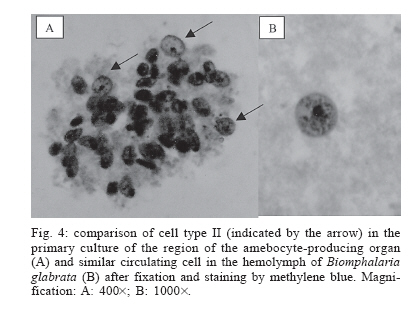
Schistosomiasis is a parasitic infection caused by the digenetic trematode Schistosoma mansoni Sambom, 1907. The disease is prevalent in 54 countries and territories of Africa, the Caribbean, the Mediterranean, and South America. It is estimated that 130 million people are infected with this species of parasite (Chitsulo et al. 2000). Although several advances have been achieved in the control of the schistosomiasis morbidity, mainly due to the development of new low toxicity and highly effective drugs against the parasite, it is still an endemic disease, whose control and/or eradication is very difficult in the wide regions where it occurs. The affected patients become debilitated, thus impairing the socio-economic development of the affected countries. In Brazil, it is estimated that eight million people are infected, whereas thirty million are at high risk of infection, since they live in hyper endemic areas (Katz 1997). The human infection starts with the penetration of the skin by larval forms of S. mansoni living in infected fresh-waters, where the parasite utilizes snails of the genus Biomphalaria Preston 1910 (Mollusca: Pulmonata, Pla-norbidae) as intermediate hosts. In Brazil, three species of Biomphalaria are considered as hosts for schistosomiasis mansoni, in this order of importance: B. glabrata, B. tenagophila, and B. straminea. In the S. mansoni-Biomphalaria interaction, besides the stringent physiological and biochemical compatibility between host and parasite, the success or failure of the infection will be strongly determined by the efficacy of the internal defense system of the snail. When in contact with the infective agent, the endogenous defense mechanisms of the mollusk are immediately activated, mediated by circulating effectors cells (hemocytes) and by soluble factors which are present in the hemolymph. Among these factors, lectins with specific carbohydrate-binding capacities play an important role by immobilizing foreign objects by agglutination, promoting phagocytes by the hemocytes or even acting as cytophilic receptors for non-self recognition (Van der Knaap & Loker 1990). The hemocytes of Biomphalaria are capable of phagocytosis, encapsulation and, finally, destruction of the infective agents. Both morphologically and functionally, these cells resemble cells of the vertebrate monocyte-macrophage series (Jeong et al. 1983, Bezerra et al. 2003). Experimental evidence has been accumulated assigning the production and storage of hemocytes to the hematopoietic organ, a structure that is located on the wall of the pericardium and posterior epithelium of the mollusk mantle. That organ, also known as amebocyte-producing organ or APO, is composed of a small quantity of primary ameboblasts that rest on a loose reticulum, which is formed by extensions of smooth muscles and fibroblasts (Jeong et al. 1983). In a recent study, Sullivan et al. (2004) reported an increase in the frequency of cell mitosis in the APO of B. glabrata following the inoculation of miracidia or cercariae antigens of S. mansoni. The mitotic ratio was intensified in Biomphalaria strains that were naturally resistant to infection by S. mansoni. Furthermore, experimental transplantation of several organs from B. glabrata specimens from a strain partially resistant to S. mansoni infection to individuals from a susceptible strain, demonstrated that the APO was unique in its ability of transfering resistance to the receptors (Sullivan & Spence 1994). More recently, Barbosa et al. (2006) transformed susceptible strains of B. tenagophila into resistant ones by transplantation of the APO from an absolute resistant strain (Taim, RS, Brazil) of the same mollusk species. The transplanted recipients became completely resistant to S. mansoni infection. The present study describes, for the first time, the primary culture of the APO region of B. glabrata. The availability of such an in vitro model may provide a useful tool for studying the biology and development of the internal defense system of these snails, as well as a better understanding of the mechanisms of susceptibility or resistance to S. mansoni in different Biomphalaria strains. This approach may suggest, perhaps, new alternatives for the control of human schistosomiasis. MATERIALS AND METHODS Materials - Mebendazole (100 mg/pellet) is a pharmaceutical product of the Ezequiel Dias Foundation. The fetal bovine serum (FBS) was processed in our laboratory from a crude stock. CMRL 1415 culture medium was prepared from individual components (Morton 1970) whereas Ham's F-10 (N 6635), Ham's F-12 (N 6760) and RPMI-1640 (R 6504) culture media were prepared according to the manufacturer's instructions (Sigma Co.). Multi-well plates were purchased from Costar Co., and poly-L-lysine (P 9155), sodium selenite (S 9133), putrescine (P 5780), pro-gesterone (P 6149), and epidermal growth factor (E 1257) were from Sigma. All the solutions, plastic, glassware and surgical tools used were previously sterilized. All other reagents were of the best grade available. Mollusks - Two hundred B. glabrata snails, laboratory reared in aquaria at the Research Center René-Rachou- Fiocruz, were used. The specimens were distributed in groups according to the diameter of their typical planorbid shells, a parameter commonly utilized to estimate the animal's age. Individuals with shell diameters between 7 to 40 mm were tested. The mollusks were kept in beakers with 250 ml of bi-distilled water containing 2.5 mg/ml mebendazole, a general anti-helminthic drug, for 48 h, until their brownish faeces turned to whitish. The intact shell of the animals was then cleaned with 70% alcohol and transferred to a laminar-flow cabinet for dissection under aseptic conditions. Primary cultures - The snail's shells were opened by means of scissors. The soft parts were separated with surgical tweezers and placed in a Petri's dish. The APO region was removed according to the procedure described by Sullivan (1990) and placed in 0.5 ml of PBS for a few seconds in order to prevent desiccation. The APO region was then rapidly sliced into small fragments, which were placed into 12- or 24-well plates containing 500 µl or 250 µl of culture medium, respectively. After adhesion of the tissue explants, 24 h later, the volumes of culture medium per well were doubled by addition of fresh medium. The plates were incubated at fixed temperatures of 15 or 20ºC. A control culture was established with the ovotestis of Biom-phalaria. The organ was removed and cultured under exactly the same conditions described for the APO region. The culture media tested were Ham's F-10, Ham's F-12, CMRL 1415, and RPMI-1640, containing 10% fetal bovine serum, 4 mg/ml fungizon and 10 mg/ml gentamicin. The collagens of South American rattlesnake (Crotalus durissus terrificus) and rat's tail (Duarte et al. 1999), poly-L-lysine and 3% agar were tested as cell adhesion helpers. Poly-L-lysine was used according to the manu-facturer's instructions. Growth promoters and differentiation inducers for vertebrate cells, such as insulin (1.25 µg/ml), sodium selenite (30 ng/ml), putrescine (0.3 µg/ml), transferrin (5 µg/ml), and EGF (0.1 ng/ml), were also tested. Viability of the primary cell cultures was determined using the trypan blue dye exclusion method and further counting of viable and non-viable cells using a hemocytometer. Cells from the hemolymph of Biomphalaria - The hemolymph of B. glabrata snails (about 200 µl/specimen) was removed from the hepato-pancreas region by means of a syringe with an insulin needle and pooled together. After enzymatic digestion with the same volume of 0.025% trypsin-0.02 MEDTA for 10 min, at room temperature, to help in cell dispersion, the solution was centrifuged at 3000 rpm for 2 min. The cell pellet was suspended in 200 µl PBS and immediately stained with methylene blue. Cells staining by methylene blue - Slides were prepared with cells from the primary culture or from the hemolymph pool. In either case, the cells were fixed in 4% paraformaldeyde for 1 h, washed once with PBS and centrifuged at 2000 rpm. The pellet was suspended in 1% methylene blue in distilled water and gently dropped in slides. After mild flame heating, the slides were covered and examined under the microscope. All the previous steps were performed at room temperature. RESULTS Cell culture conditions - The cell culture
experiments were carried out with mollusks whose shell diameters
varied between 12 and 18 mm. Among the culture media tested, the
best results were obtained with CMRL 1415 supplemented with insulin,
transferrin, sodium selenite, putrescine, EGF, and FBS. Under this
condition, the cells viability was 75% in average, after one week
in culture. From the second week onwards, signs of cell death were
progressively observed, such as the presence of granulations in
the cytoplasm and swelling followed by lysis. The cultures, then,
were maintained for a maximum period of two weeks. In the absence
of FBS there was nor cell migration from the explants of the APO
region neither cell growth, whereas in the cultures without EGF
the growing cells consisted mainly of fibroblasts. No significant
differences were observed in the growth pattern in the presence
of adhesion substrates. The best temperature for cell growth was
15ºC, which also favored the migration of cells from the APO
explants. The migration was impaired when the incubation temperature
was raised to 20ºC, followed by death of every cell present
within the first 24 h. After 1 week at 15ºC, the temperature
increase to 20ºC led to a rapid decrease in cell viability
from 75 to 5% within Cell types in culture - The cells migrated effortlessly from the explants of the APO region, with no need of any enzymatic treatment to assist cell dispersion in culture. A number of cells already dispersed in the medium could be observed starting from the very first day of culture (Fig. 1). Three cell populations or types morphologically distinct, named I to III, grew in suspension in the cultures of the APO region (Fig. 2). These closely resembled two cell types known to occur in the hemolymph of Biomphalaria. Type I cells (Fig. 2A,B), which comprised the predominant population in culture, were rounded with a high nucleus to cytoplasm ratio, morphologically similar to hyalinocytes of the hemolymph (Van der Knaap & Loker 1990). The cell population type II displayed relatively more cytoplasm than type I (Fig. 2A,B) in accordance with the description of granulocytes from the hemolymph (Van der Knaap & Loker 1990). Extended pseudopods, previously reported as an important ability of granulocytes, were also observed in the cultures (Fig. 2C). However, it was not possible to identify the cell type they came from. The last cell population in culture, named III, was characteristically light-refringent under phase-contrast microscopy, with no defined nuclei (Fig. 2B). This population was much less abundant than the previous two and, as far as we know, has not been reported before in the molluskan hemolymph. Interestingly, atypical cell division, with nuclei duplication without concomitant cytokinesis, was observed in the cultures after the cells migrated from the explants of the APO region (Fig. 2D). In the primary culture of the ovotestis (control) the cells grew in clusters, strongly adhered to the substrate (Fig. 3). A single cell type was observed in the hemolymph of B. glabrata, the morphological and staining pattern of which resembled the cells from population II in the primary cultures of the molluskan APO region (Fig. 4A,B, respectively). DISCUSSION The first trials of tissue culturing of the APO region were performed with B. glabrata specimens with shell diameters between 6 and 40 mm. Handling and dissection of younger specimens (shell diameters below 12 mm) were hampered by their small size. The first difficulty encountered with much older specimens (shell diameters above 20 mm) was the high rate of natural contamination of the mollusks by a variety of endoparasites. About 50% of the specimens had to be discarded in the very first days in the laboratory due to the persistence of high levels of parasitism even after mebendazol treatment. Individuals with shell diameters between 12 and 18 mm were the most responsive to the drug and with a reduction of up to 90% in the contamination rate. These specimens were, then, used in the experiments. The number of circulating hemocytes was previously described to be highly variable in Biomphalaria and mainly dependent on parameters such as the method and local of hemolymph extraction besides the physiological conditions of the snail (Sminia 1981). Similarly, there is no agreement in the literature on the number and relative frequency of the cell types most commonly found in the hemolymph of Biomphalaria. The hemocytes do not behave as a homogeneous population and morphological/biochemical heterogeneity among them have been described by several authors. Sminia et al. (1983) reported that gastropods, in general, possess a unique type of hemocyte, which suffers morphological changes during cell maturation. The youngest cells are rounded and highly active for mitoses, whereas the mature ones display cyto-plasmatic indentations, low mitotic and high phagocytic activities. The presence of two sub-populations of hemocytes mainly differing in size, phagocytic activity and ability to emit pseudopods, was demonstrated by others (Cheng & Auld 1977, Van der Knaap & Loker 1990, Barraco et al. 1993, Johnston & Yoshino 2001). On the other hand, three hemocyte cell types were described by Matricon-Gondran and Letocart (1999). Among these, one cell type was found to share typical properties of mammalian macrophages, such as high phagocytosis activity, resistance to elevated doses of gamma radiation and the presence of inner lysosomal structures (Bezerra et al. 2003). An even higher number of hemocyte types were described by Joky et al. (1983). Up to four cell types were proposed based not only on morphological characteristics, but also on their ability of expressing different surface lectin receptors. Independently of the inconsistency in the definition and characterization of types, there is a consensus that the hemocytes and their products represent the basic elements of the innate defense system of Biom-phalaria, responsible for the phagocytosis or encapsulation of small size (bacteria, fungi and protozoa) or large size (larvae of helminthes) pathogens, respectively (Adema & Loker 1997). A significant number of authors have reported the presence of two sub-populations of hemocytes, the so called granulocytes and hyalinocytes (Cheng & Auld 1977, Van der Knaap & Loker 1990, Barraco et al. 1993). With 5 to 8 µm in diameter and a large nucleus to cytoplasm ratio, the hyalinocytes have a circular contour when in contact with polished glass surfaces, little tendency to form pseudopods and scarce lysosomal structure. On the other hand, the granulocytes have diameters between 7 and 11 µm with more cytoplasm. They emit pseudopods, are active in phagocytosis and represent more than 90% of the circulating cells (Barraco et al. 1993). This observation explains why a single cell population, was observed in the hemolymph of B. glabrata, most probably corresponding to this major population (granulocytes). Two cell types, similar to the two hemocyte populations previously described based mainly on their nucleus to cytoplasm ratio, were observed in the cultures of the APO region of B. glabrata. These cultures, unfortunately, were limited to short-term ones. Indeed, the difficulty of obtaining hemocytes growing in vitro was pointed out before (Lynn 2001). One possible reason for that could be the low proliferation rate of the cells (Lebel et al. 1996). The morphology of the cells that grew in the primary culture of the ovotestis (control) was totally distinct of the cell populations observed in the culture of the APO region. These data suggest that the origin of hemocytes is possibly restricted to specific organs/regions, such as the APO of Biomphalaria. Recently, based on morphological data, Souza and Andrade (2006) suggested a multicentric origin of the hemocytes, starting at the endothelium of vascular spaces. Due to the limited data available for the culturing of hematopoietic tissues of invertebrates, our attempts were based on studies in vertebrates, in which the proliferation/differentiation rates are known to be controlled by hormones, growth factors, cytokines, surface receptors, and genes (Watt & Hogan 2000). Cultures of invertebrate tissues have been described as being insensitive to many of these factors that affect vertebrate systems (Slack 2000, Watt & Hogan 2000). However, Lebel et al. (1996) reported a positive effect in invertebrate cell cultured in the presence of fetal bovine serum and epidermal growth factor. Our data are in accordance to the aforementioned results. The present work describes for the first time the successful primary culture of the APO region of B. glabrata. Further work is in progress to isolate the cell populations obtained so far in the primary cultures with the aim of gaining a more detailed morphological and functional characterization. ACKNOWLEDGEMENTS To Ms MS Hermógenes and AC Valentim for excellent technical assistance. To Ms DMS Reis, DM Repetição, and SSF Teixeira, technicians in charge of the maintenance of the Biom-phalaria aquaria at the CPqRR-Fiocruz. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:Photo images[oc06108f4.jpg] [oc06108f3.jpg] [oc06108f1.jpg] [oc06108f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}