 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101, No. 6, September ,2006, pp. 693-696 Giardia duodenalis: analysis of secreted proteases upon trophozoite-epithelial cell interaction in vitro Guadalupe Beatriz Rodríguez-Fuentes/++, Roberto Cedillo-Rivera*, Rocío Fonseca-Liñán**, Raúl Argüello-García**, Onofre Muñoz, Guadalupe Ortega-Pierres**, Lilián Yépez-Mulia/+ Unidad de Investigación Médica en
Enfermedades Infecciosas y Parasitarias, Hospital de Pediatría,
Centro Médico Nacional Siglo XXI, Instituto Mexicano del
Seguro Social-Universidad Autónoma de Yucatán, Mérida,
Yuc., México *Unidad Interinstitucional de Investigación
en Salud, Instituto Mexicano del Seguro Social, México, D.F.,
México **Departamento de Genética y Biología
Molecular, Centro de Investigación y de Estudios Avanzados,
IPN, México, D.F., México
Financial support: Conacyt project 4315P-M9608, Programas de Apoyo a la Investigación Médica, Coordinación de Inves-tigación Médica, Mexico, project F0038-R5 Received 23 March 2006 Code Number: oc06118
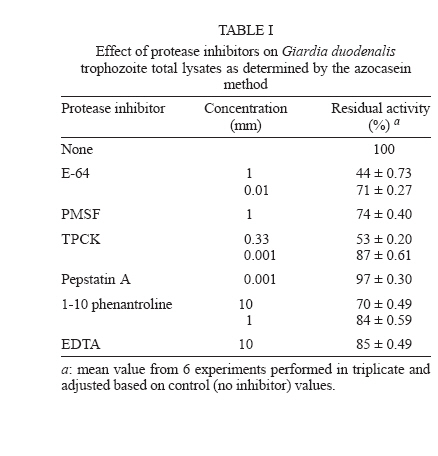
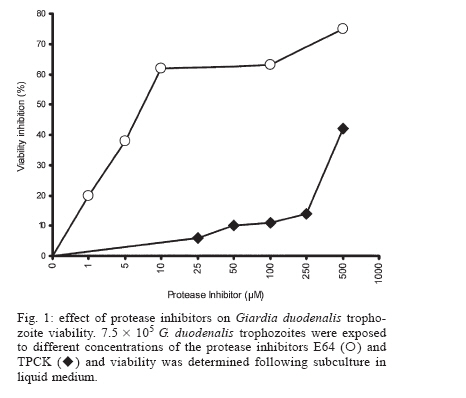
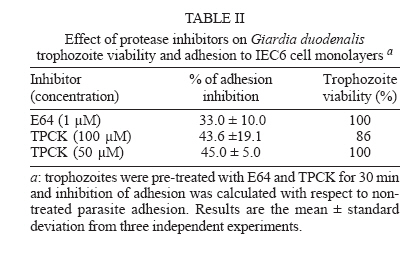
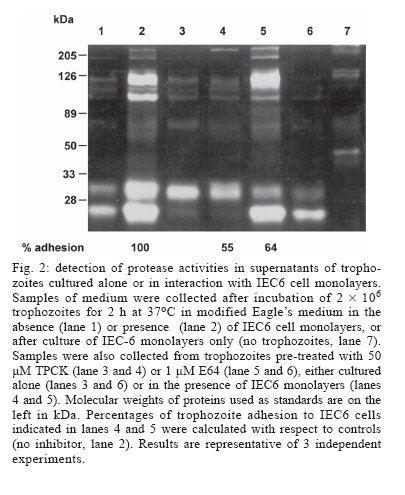
Protease secretion by Giardia duodenalis trophozoites upon interaction with epithelial cells and its association with the parasite adhesion was studied in co-cultures of parasites with IEC6 epithelial cell monolayers in the presence or absence of protease inhibitors. Proteolytic activity in supernatants from trophozoites was enhanced when they were co-cultured with IEC6 cells. This activity was strongly inhibited by pre-incubation of live trophozoites with E-64 and TPCK and a concomitant inhibition of parasite adhesion to IEC6 cells was observed. These data suggest that trophozoites secrete cysteine-type proteases that play a role in the adhesion of G. duodenalis to epithelial cells. Key words: Giardia duodenalis - protease secretion - adhesion Giardiasis caused by the flagellated, intestinal protozoan parasite Giardia duodenalis (syn. G. lamblia, G. intestinalis) manifests as acute and chronic diarrhoea in communities of both developing and developed countries. It is also a leading cause of defined waterborne diarrhoea worldwide (Adam 1991). Infection results from ingestion of cysts, most commonly from faecal contamination of water and food whereas community-wide outbreaks result from contaminated water supplies. After excystation in the upper small intestine, adhesion of trophozoites to epithelial intestinal cells represents the first step in the pathogenesis of the disease. The use of IEC6, an epithelial cell line derived from rat small intestine, is a well validated model in which the inhibitory effect of both actin inhibitors and chelating agents on the attachment of G. duodenalis trophozoites has been previously demonstrated (McCabe et al. 1991). It is also well recognized that the adhesion process is a multi-factorial event that involves a suction force of the adhesive disk (Knaippe 1990), a mechanical process related to contractile proteins of the adhesive disk and the ventrolateral flange (Chávez et al. 1995) and surface molecules that allow the binding of trophozoites to receptors to host epithelial cells (Pegado & Souza 1994). It has been postulated that proteases may be involved in this process. Several studies have demonstrated protease activity in total extracts of Giardia trophozoites or in parasite vacuoles (Feely & Dyer 1987, Hare et al. 1989) including the Portland 1 (P1) strain cultured in vitro, with the predominance of cysteine, serine, and aspartic protease activities (Williams & Coombs 1995). It has also been shown that cysteine proteases of P1 strain degraded protein substrates such as gelatin, collagen, albumin, and azocasein while serine proteases were mainly active against haemoglobin (Coradi & Guimarães 2006). In addition, the presence of cysteine protease activities in excretory/secretory (E/S) products of P1 strain has also been reported (Jiménez et al. 2000). These E/S products are associated with mucosal injuries observed in murine giardiasis (Jiménez et al. 2004). However the possible relation between protease secretion and the adhesion of G. duodenalis to epithelial cells has not been tested. The aim of this study was to identify proteases secreted by Giardia upon interaction with the epithelial cell line IEC6 and to determine their possible role in parasite adhesion. Trophozoites of WB strain (ATCC # 30957) were cultured in TYI-S-33 medium supplemented with 10% calf serum (Hyclone, Logan UT, US), in the presence of 2 mM glutamine and antibiotics (10 µg/ml streptomycin, 10,000 UI/ml penicillin, and 2.5 µg/ml amphotericin B). The protease activity of trophozoite extracts was determined by the method of Beynon and Bond (1994). To measure inhibition of protease activity 100 µg of total extracts in 100 µl were pre-incubated for 30 min at 37ºC with a range of concentrations of protease inhibitors: trans-epoxysuccinyl-L-leucylamido (4-guanidino)-butane (E64) to identify cysteine proteases; L-1-chloro-3-[4-tosylamido]-4-phenyl-2-butanone (TPCK) for serine and cysteine proteases; phenylmethanesulphonyl fluoride (PMSF) for serine proteases; pepstatin A for aspartic proteases, and ortho-phenanthroline (1,10- phenanthroline) and EDTA, for metallo proteases. All protease inhibitors were purchased from Sigma, St. Louis MO, US. 250 µl of 0.2% azocasein (w/v) in 0.1 M acetate buffer pH 5.5 were added to the mixture and incubated for 1 h at 37°C. The reaction was stopped by the addition of 10% trichloroacetic acid (v/v) and then centrifuged for 15 min at 800 ´ g. The absorbance of the supernatants was determined at 366 nm to measure azocasein proteolysis. The effect of protease inhibitors on the proteolytic activity present on total lysates is shown in Table I. The highest percentages of inhibition of azocasein proteolysis were obtained when 1 mM E64 (56%), 0.33 mM TPCK (47%), 10 mM 1-10 Phenanthroline (30%) and 1 mM PMSF (26%) were used. In the case of Pepstatin A (from 1 mM to 100 µM) only 3 to 5% inhibition was obtained. These data are consistent with a predominance of cysteine-type proteases in lysates from G. duodenalis trophozoites and are in agreement with similar assays carried out in Portland 1 strain lysates (Williams & Coombs 1995, Coradi & Guimarães 2006). Because of this and the likelihood that intracellular proteases give rise to mature extracellular proteases, E64 and TPCK were selected to evaluate the role of cysteine proteases in the adhesion of trophozoites to IEC6 cells. As a prerequisite for adhesion assays, we determined the parasite viability at different concentrations of these protease inhibitors (Fig. 1) using subculture in liquid medium. Briefly, trophozoites (7.5 ´ 105) were incubated with the inhibitors for 30 min at 37oC, washed twice with PBS and subcultured in fresh medium for 48 h at 37oC. The cells were counted and their viability was determined by interpolation in a control curve obtained with different inocula of untreated trophozoites (Argüello-García et al. 2004), and the experimental lethal concentrations at 50 and 20% (LC50 and LC20 respectively) were also calculated. The data obtained showed that E64 was a potent inhibitor with a LC50 value of 7.47 µM. In comparison the LC50 for TPCK was 663.5 µM. These calculations were in good agreement with results in experiments using these LC50 values. For adhesion assays trophozoites (2 ´ 106/well) were co-cultured with confluent monolayers (485 ´ 103 cells/well) of the epithelial cell line IEC6 (ATCC accession CRL-1592) for 2 h at 37°C in a 5% (v/v) CO2 atmosphere in modified Eagle's medium (Sigma) without serum. These conditions allow a maximum (~70%) of parasite adhesion (McCabe et al. 1991). Trophozoite adhesion was quantified by thymidine-labelling of parasites as described previously (Bermúdez-Cruz et al. 2004). To determine the effect of cysteine protease inhibitors on parasite adhesion, 2 ´ 106 trophozoites were pre-incubated with 1 µM E64 or 50 µM TPCK for 30 min, i.e., at concentrations below the LC20 values for E64 and TPCK (1.05 µM and 234.94 µM respectively). After incubation, parasite viability was confirmed by flagellar motility. Trophozoites incubated with IEC6 cells without protease inhibitor treatment were included as adhesion controls. The mean viability observed post-treatment was 100% for both compounds (Table II). Pre-treatment with either inhibitor decreased trophozoite adhesion and differences between the two inhibitors were not statistically significant (P > 0.05, Table II). However, TPCK consistently showed the highest levels of inhibition of adhesion (~ 46%) while E64 inhibited the parasite adhesion by ~ 33%. Therefore, parasites pre-treated with 50 µM TPCK or 1 µM E64 were used to further analyze parasite secreted proteases upon interaction with epithelial cell monolayers. Supernatants from the adhesion assays were collected, filtered through a 0.22 µm membrane and concentrated by ultrafiltration with a 10 kDa exclusion pore (Amicon). Supernatants from trophozoites pre-treated or not with protease inhibitors and cultured alone (in the absence of IEC6 monolayers) were also included. Electrophoretic analysis of proteolytic activity (30 µg protein/well) in these supernatants was carried out at 140 V, 2 h, 4ºC in 10% SDS-polyacrylamide gels (w/v) co-polymerized with 0.2% gelatin (w/v) (Sigma). After electrophoresis, gels were incubated for 30 min at 37ºC with 2.5% Triton X-100 , washed twice in distilled water and incubated in 0.1M sodium acetate buffer at pH 5.5 for 6 h. Gels were stained with Coomassie blue (BioRad) and analyzed by densitometry using a Foto-dyneTM image analyzer integrated with a Macintosh computer. When the supernatant of trophozoites cultured alone was analyzed, 7 bands of protease activity were detected representing MWs of 198, 135, 120, 107, 73, 30, and 22 kDa (Fig. 2, lane 1). Among these, the 30 and 22 kDa proteases were the most active. After the interaction of WB trophozoites with IEC6 cells, the activity of the 135, 107, 30, and 22 kDa proteases increased by 182, 150, 202, and 250%, respectively (Fig. 2, lane 2). A similar enhancement of protease secretion by trophozoites was observed when they were co-cultured with MCDK monolayers (data not shown). However in supernatants of WB trophozoites pre-incubated with TPCK and cultured alone (Fig. 2, lane 3) or with IEC6 (Fig.2, lane 4), proteolytic activity (except for the 30 kDa protease band) was significantly inhibited. On the other hand, in the supernatant of trophozoites pre-incubated with E64 and cultured alone, most of the proteolytic activity was inhibited or markedly reduced, except for the 22 kDa protease band (Fig. 2, lane 6). When trophozoites pre-incubated with E64 were co-incubated with IEC6 cells (Fig. 2, lane 5), an increase in proteolytic activity was observed for proteases of 198 kDa (105%), 135 kDa (200%), 107 (90%), and 22 kDa (120%). The proteolytic activity of the 30 kDa protease band increased to a lesser extent (50%). Also an additional protease with MW of 180 kDa was observed. Supernatants of IEC6 cultured alone revealed proteases of 213, 194, 149, 139, 73, 48, and 43 kDa with minor activities (Fig. 2, lane 7). Only the 73 kDa protease band was common to all supernatants. Thus the proteases of 198, 135, 120, 107, 30 and 22 kDa detected in supernatants from trophozoites co-cultured with IEC6 cells or from parasites cultured alone were parasite-derived. Although cysteine proteases of 95 kDa, 38 kDa, and 35 kDa from WB strain have been purified (Parenti 1989, Werries et al. 1991) this is, to our knowledge, the first time that proteases from total lysates of WB strain have been analyzed. These activities corresponded mainly with cysteine, serine and metallo proteases. Although Williams and Coombs (1995) reported multiple protease activities in the P1 strain, predominantly of cysteine, serine and aspartic nature, in our study the aspartic inhibitor pepstatin A had no effect on the proteolytic activity of WB total lysates, even at 100 µM. This is supported by similar observations in previous reports with other substrates and the P1 strain (Hare et al. 1989, Parenti 1989). Mining of the Giardia Genome Project based on a clone (A6) from WB strain (http://www.embl.edu/Giardia) revealed at least 7 well characterized genes for giardial cysteine proteases while the deduced open reading frames for other putative proteases predicted only serine and metallo but not aspartic-type proteases. Our findings are also consistent with the detection of proteolytic activity in E/S products from trophozoites of the P1 strain (Jiménez et al. 2000, 2004). However an enhancement of protease secretion during trophozoites-IEC6 interaction has not been previously reported. Interestingly when G. duodenalis trophozoites were cultured in the presence of a cytosol-enriched fraction from MDCK cells, an increase in protease secretion was observed while membrane-enriched fractions were not effective (data not shown). We are currently characterizing epithelial cell molecule(s) responsible for enhancing parasite protease secretion. The association between the inhibition of adhesion of G. duodenalis trophozoites treated with TPCK or E64 to IEC6 cells and the concomitant inhibition of major protease activities in supernatants of this interaction supports the notion that protease secretion plays a role in the adhesion of the parasite to epithelial cells as suggested for other protozoa (Arroyo & Alderete 1995, García-Rivera et al. 1999). Several mechanisms have been proposed to explain the role of parasite proteases in the adhesion process. These include the unmasking of putative receptors on the membrane of either epithelial cells or trophozoites, as suggested for Trichomonas vaginalis (Arroyo & Al-derete 1995), or the association of cysteine proteases with adhesins as in Entamoeba histolytica (García-Rivera et al. 1999). Alternatively, in vivo proteases may play a role in the degradation of mucin, thus facilitating either adhesion of parasites to target cells or parasite motility (Paget & James 1994). The precise mechanism by which secreted proteases of G. duodenalis participate in the adhesion process certainly requires further studies. ACKNOWLEDGEMENTS To Jacqui Upcroft for critically reviewing this manuscript and for valuable suggestions. To Rosa María Bermúdez for comments on the manuscript. To Arturo PérezTaylor, René López, and Blanca Herrera for technical assistance. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:Photo images[oc06118f2.jpg] [oc06118t2.jpg] [oc06118t1.jpg] [oc06118f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}