 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101, No. 7, November ,2006, pp. 715-719 Identification and molecular characterization of Van A-type vancomycin-resistant Enterococcus faecalis in Northeast of Brazil Marinalda Anselmo Vilela, Sandra Lavareda de Souza*, Izabel Cristina Vanzato Palazzo**, Joseane Cristina Ferreira**, Marcos Antonio de Morais Jr*, Ana Lúcia da Costa Darini**, Márcia Maria Camargo de Morais/+ Departamento de
Patologia, Instituto de Ciências Biológicas *Departamento de
Genética e Laboratório de Imunopatologia Keizo Asami, Universidade
Federal de Pernambuco, Rua Arnóbio Marques 310, 50100-130 Recife,
PE, Brasil +Corresponding author: camargo@icb.upe.br Financial support: Facepe (PPP-006/2003), Pró-Reitoria de Pesquisa, USP (2004.1.25515.1.8) Received 21 Februarey
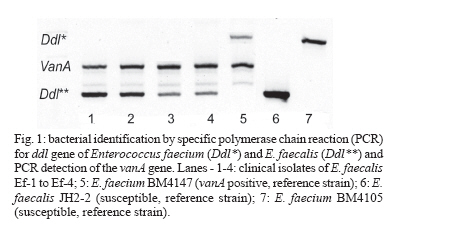
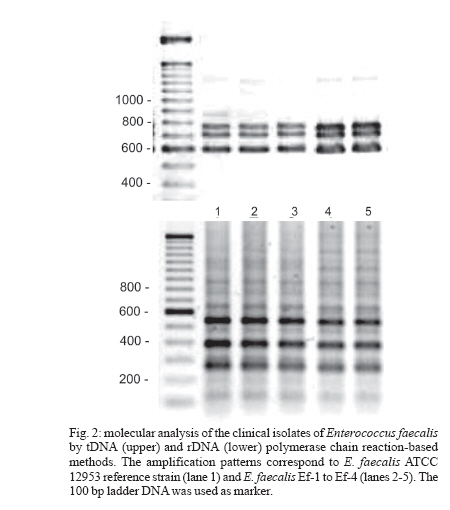
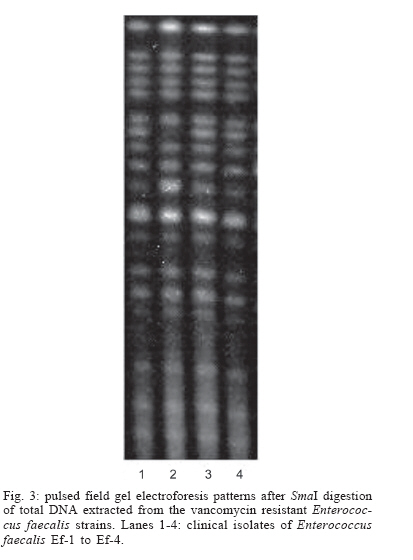
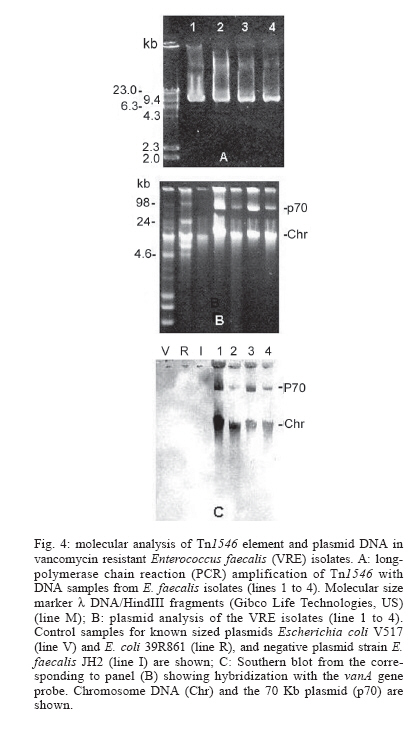
2006 Code Number: oc06120 The isolation of vancomycin resistant enterococci (VRE) in Brazil has rapidly increased, following the world wide tendency. We report in the present study the first isolation of vancomycin resistant Enterococcus faecalis (VRE) in the Northeast of Brazil. The four VRE isolates were characterized for antimicrobial susceptibility, genotypic typing by macro restriction of chromosomal DNA followed by pulsed-field gel electrophoresis and for characterization of the Tn1546-like element and plasmid contents. The isolates showed resistance to multiple antibiotics and a single genotype profile, suggesting the dissemination of a single clone among the patients. Tn1546 associated to genetic elements as plasmids shows the importance of infection control measures to avoid the spreading of glycopetide resistance by conjugative transfer of VanA elements. Key words: vancomycin resistance - Enterococcus faecalis - transposable elements Enterococci are opportunistic microorganisms, which have emerged as cause of nosocomial infection, usually in patients of long-term hospitalization (Furtado et al. 2005). In the last years, reports on vancomycin-resistant enterococci (VRE) nosocomial infections have rapidly increased in many countries worldwide, and this pathogen is now being recognized of clinical relevance. High resistance against vancomycin and teicoplanin results from the altered affinity of this glycopeptide for its natural target, which occurs due to acquisition of Tn-1546 carrying vanRSHAX genes and elements closely related to it (Courvalin 2006). The transposon encodes the VanH dehydrogenase that reduces pyruvate to D-lac, and the VanA ligase that catalyzes the formation of an ester bond between D-ala and D-lac. The resulting D-ala-D-lac depsipeptide replaces the D-ala-D-ala dipeptide in peptidoglycan synthesis, a substitution that decreases the affinity of the molecule for the antibiotic glycopeptides (Courvalin 2006). In Brazil, the first VRE isolated occurred in 1996 (Dalla Costa et al. 1998) and many cases have been described thereafter (Zanella at al. 1999, 2003, Dalla Costa et al. 2000, Cereda et al. 2001, Reis et al. 2001, Almeida et al. 2004, Camargo et al. 2004, Furtado et al. 2005). However, those studies were restricted to the Southeast, Central, and South regions of Brazil. Nevertheless, the rapid spreading suggested that novel cases should soon be detected in Northern and North regions as well. In this report we describe the isolation of four VRE in Recife, state of Pernambuco, Northeast of Brazil, which seems to be the first report in this region. MATERIALS AND METHODS Bacterial strains and clinical features - The VRE isolates (n = 4) studied were isolated in 2004, from patients of a teaching university hospital (Hospital Universitário Oswaldo Cruz HUOC), located in the city of Recife, Pernambuco, Northeast of Brazil. The hospital has 342 beds and belongs to the State University of Pernambuco. The bacteria were stored at _80ºC. The VRE isolates were collected from four patients at post-surgical period, after being submitted to gastroplastic surgery (one patient), exploratory laparatomy (one patient), and liver transplantation (two patients). They remained in the hospital from 41 to 60 days at intensive care unit (ICU), sharing 10 days all together at the same hospital sector. Two patients died, one of them presenting positive blood culture for VRE. Isolates were presumptively identified as enterococci by colonial morphology, Gram's stain, the absence of catalase production, the presence of pyrrolidonylarylamidase by hydrolysis of L-pyrrolidonil-b-naphthylamide (Becton, Dickinson & Co., Franklin Lakes, NJ), tolerance to 6.5% sodium chloride, ability to grow at 10ºC and 45ºC, growth on bile-esculin agar, with esculin hydrolysis, and growth on tellurite agar. Susceptibility testing - Screening for antibiotic resistance was performed by disk diffusion method on Mueller-Hinton agar according to "Clinical and Laboratory Standard Institute" guidelines (CLSI 2005) for the following antimicrobial agents: chloramphenicol, penicillin, erythromycin, vancomycin, teicoplanin, ciprofloxacin, gen-tamycin, streptomycin, ampicillin, linezolide, and tetracycline. The minimum inhibitory concentration (MIC) for vancomycin and teicoplanin was determined by Etest. The control strains used in this work were Enterococcus casseliflavus NCTC 12361, E. faecium NCTC 7171, E. faecalis NCTC 775, E. gallinarum NCTC 12359, E. faecium BM 4147 vanA genotype, E. faecium BM4105, E. faecalis JH2-2, and S. aureus ATCC 25923 for MIC assays. Molecular identification of VRE isolates - Bacterial DNA was extracted by rapid alkaline lysis method and used as template in amplification reactions according to Depardieu et al. (2004). Briefly, the identification of E. faecalis isolates was done by the amplification of the 475-bp fragment corresponding to the ddlE. faecalis gene using the primers DD13(+) (5'-CACCTGAAGAAACAGGC-3') and DD3-22(-) (5'- ATGGCTACTTCAATTTCACG-3'). Control amplification was performed using the E. faecalis JH2-2 reference strain. Additionally, DNA of the E. faecium reference strains BM4105 and BM4147 were amplified with the primers FAC1-1(+) (5'-GAGTAAATCACTGAACGA-3') and FAC2-1(-) (5'-CGCTGATGGTATCGATTCAT-3') to produce the 1091-bp species-specific fragment corresponding to the ddlE. faecium gene. Detection of the vanA gene - The bacterial DNA samples were used for the detection of vanA gene by the use of the primers EA1(+) (5'-GGGAAAACGACAATTGC-3') and EA2(-) (5'-GTACAATGCGGCCGTTA-3') according to Depardieu et al. (2004). Genotyping analysis - The bacterial isolates were submitted to two different approaches: polymerase chain reaction (PCR)-fingerprinting and pulsed-field gel electrophoresis (PFGE). For PCR-fingerprinting, total genomic DNA of the four VRE isolates was obtained by alkaline lysis method (Sambrook et al. 1989) and used as template for the amplification of ITS-rDNA sequences with the primers KT03(16S) (5'-TTGTACACACCGCCCGTCA-3') and KT04(23S) (5'- GGTACCTTAGATTGTTTCAGTTC-3') according to Kostman et al. (1992). Additionally, PCR analysis of the tRNA-encoding sequences used the primers T5A (5'- AGTCCGGTGCTCTAACCAACTGAC) and T3B (5'- AGGTCGCGGGTTCGAATCC-3') according to Welsh and McClelland (1991). For both markers E. faecalis ATCC 12953 was used as reference strain. The resulting amplification fragments were separated on 1.2% agarose gel at 7 V.cm-1 in 0.5 X TBE buffer. PFGE analyses of SmaI-digested bacterial DNA was performed in Gene Navigator apparatus (Amersham, Upsalla, Sweden) at 180V for 25 h, at 7ºC, as described previously by Campanile et al. (2003). The equipment was adjusted for pulses of 20 s for 10 h, 8 s for 10 h, and 3 s for 5 h. DNA banding patterns were visualized at UV light after staining with ethidium bromide. The isolates clonality was judged using previously described criteria (Tenover et al. 1995) by comparing visually the banding patterns of samples that have run together in the same gel. Amplification and characterization of Tn1546-like elements - Amplification of Tn1546- like elements and vanRSHAX regions was performed by Long-PCR (L-PCR) according to the method described by Palepou et al. (1998). Briefly, total genomic DNA of the strains was extracted as described by Pitcher et al. (1989) and used as templates for PCR with the single primer P1 (5'- GGAA AATGCGGATTTACAACGCTAAG-3'), complementary to the transposon-flanking inverted repeats, using the Expand Long Template PCR System (Roche, Germany) to produce a 10,800-bp fragment. Subsequently, the amplified Tn1546-like element was the template for L-PCR of vanRSHAX genes using primers P2 (5'- AGACAAGT CTGAGATTGACCTTGCC-3') and P3 (5'- ATATGCTT GAAACCCACTGTTTTCC-3') to produce a 4400-bp fragment. Primary PCR products were directly analyzed on 1% agarose gel or digested overnight with ClaI for the entire Tn1546-like elements or with DdeI for the vanRSHAX genes (Palepou et al. 1998). The resulting fragments were separated by electrophoresis through 2% agarose gels. Plasmid profiles
- Plasmids DNA were extracted by alkaline lysis (Birnboim and Doly
1979), and the profiles were compared after electrophoresis on 1% agarose
gels. The size of plasmids were measured by comparing their mobility with
those of plasmids of known size as the ones from E. coli V517 (NCTC
50193) and E. coli 39R861 Southern hybridization and digestion with EcoRI - Plasmids were digested with EcoRI, transferred onto nylon membrane and also hybridized with digoxigenin-labelled probe for the vanA gene. Plasmid DNA was Southern blotted onto nylon membranes (Hybond-N+, Amer-sham, Upsalla, Sweden) with a VacuGene vacuum blotting apparatus (Amersham), and the blots were hybridized with digoxigenin-labelled vanA-specific probe. This probe was prepared by PCR from genomic DNA of the strain E. faecium BM 4147 with primers and conditions previously described (Coque et al. 1998) except that the concentration of dTTP (10 mM) was reduced and digoxigenin-labelled dUTP (1 mM) was included. Hybridization was performed according to the DIG System user's guide for filter hybridization (Roche, Germany). RESULTS Bacterial identification - The isolates were confirmed for the species E. faecalis by conventional biochemical tests and subsequently by the amplification of the specie-specific 475 bp fragment of ddl gene (Fig. 1). Bacterial susceptibility - The four isolates displayed the same susceptibility profiles, showing concomitant resistance to chloramphenicol, penicillin, erythromycin, teicoplanin, ciprofloxacin, and tetracycline. It was also detected high level of aminoglicoside resistance (gentamicin and streptomycin). On the other hand, all the isolates were susceptible to ampicillin and linezolide in disk diffusion tests. MIC for vancomycin and teicoplanin was higher than 256 µg/ml. Van gene detection - All isolates harbored the vanA gene as demonstrated by PCR analysis (Fig. 1), which confirmed the phenotype of resistance to both vancomycin and teicoplanin. Genotyping analysis of chromosomal DNA - Genetic analysis by rDNA and tDNA markers showed the same pattern for all the isolates and for the reference strain (Fig. 2), which was corroborated by the identical PFGE patterns (Fig. 3). It indicates the clonality among these isolates. Detection of Tn1546-like elements and plasmids - The presence of the Tn1546-like elements harboring the vanRSHAX genes was confirmed for all VRE isolates by L-PCR method Fig. 4). Moreover, enzymatic digestion of this fragment with ClaI and DdeI showed the expected results for the complete sequence of the transposon. All results indicated the integrity of the Tn1546 in these VRE isolates. The plasmid analysis showed the presence of a 70 kb plasmid DNA for all isolates (Fig. 4b) and Southern hybridization revealed that those plasmids harbored copies of the vanA gene (Fig. 1). Positive vanA hybridization in the chromosomal DNA band in the plasmid extractions suggests a second copy of the Tn1546. In addition, Southern hybridization showed positive hybridization to the 4.3 kb EcoRI fragment of these plasmids with vanA probe (data not shown). DISCUSSION The emergence of vancomycin resistant enterococci changed the clinical relevance of enteroccocal infections through the world. In Southeast of Brazil, the isolation of such pathogens is more frequent, since many works have been reporting their rapid emergency (Dalla Costa et al. 2000, Zanella at al. 1999, 2003, Reis et al. 2001, Cereda et al. 2001, Almeida et al. 2004, Furtado et al. 2005). In the present study we describe the first isolation of a vancomycin-resistant E. faecalis in Recife, Northeast Brazil, indicating that the vancomycin resistance might be rapidly spreading through the country. Due to the phenotype of high level of vancomycin resistance, the isolates were tested for the presence of specific resistance genes (vanA and vanB) by PCR, which indicated the presence of the vanA gene. The isolates investigated showed 100% of susceptibility to ampicillin, similarly to the results reported by Reis et al. (2001). However, they were highly resistant to aminoglicosides, limiting thus the success of such associated antibiotic therapy. The concomitant resistance observed to chloramphenicol, penicillin, erythromycin, teicoplanin, ciprofloxacin, and tetracycline confirms that resistance of enterococci to multiple antibiotics is common, as also observed by Panesso et al. (2002). Linezolide was highly effective against all the isolates, indicating it as an appropriate therapeutic option. The vanA gene cluster has been reported associated to Tn1546 or related elements, which are carried by plasmids. The localization of vanA gene in a plasmid DNA and the integrity of the Tn1546 showed by VRE isolates studied are in agreement with published data (Arthur et al. 1996). Also, the integrity of Tn1546-like elements showed by Brazilian VREs studied so far (Palazzo et al. 2006) and the isolates of this study produced an important epidemiological data about the dissemination of Tn1546-like elements in Brazil. The presence of vanA gene in mobile DNA elements, such as plasmid carrying transposons, indicates that this resistance mechanism could rapidly spread by conjugation between VRE and vancomycin-sensitive enterococci (VSE) strains at appropriate intestinal environment of the patients (Fines et al. 1999), leading to spreading of glycopeptide resistance. In connection to the resistance evolution, the use of DNA-based typing methods offers high sensitivity and specificity at two levels: species identification and strain discrimination. The amplification of the specie-specific ddl gene fragment (Dutka-Malen et al. 1995) confirmed the species identification of the isolates. Additionally, PCR rDNA and tDNA genotyping succeeded in identifying the isolates at species level when comparing to reference strains, as previously described by PCR-tDNA for Enterococci (Baele et al. 2000). PFGE analysis has been long used for monitoring the dissemination of VRE clones in different countries (Suppola et al. 1999, Duckro et al. 2005, Stampone et al. 2005). Our data on PFGE strongly suggested the dissemination of a single VRE clone among the hospitalized patients in Recife. Moreover, three isolates were originated from the same hospital sector while one isolate was from a different one. The PFGE profile observed for the E. faecalis analyzed in the present study was closely related to those found in other states of Brazil (São Paulo, Paraná, Rio Grande do Sul, and Minas Gerais), indicating a clonal spread among this specie in Brazil (Palazzo et al. 2006). In conclusion, the VRE cases in Recife were result of a single clone that disseminated for different patients and hospital environments. The Tn1546 integrity and vanA gene associated to plasmids may facilitate the dissemination of vancomycin resistance among enterococci. One important implication of VRE infections is that the antibiotic therapy is now compromised. The results pointed out for the importance of the rational use of vancomycin and the execution of infection control measures as well. ACKNOWLEDGMENTS To Mr Carlos LN Cabral for his help in bacterial isolation and to Dr Patrice Courvalin and Dr Florence Depardieu for their kindly contribution to van genes detection and bacterial identification. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06120f4.jpg] [oc06120f2.jpg] [oc06120f3.jpg] [oc06120f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}