 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101, No. 7, November ,2006, pp. 755-757 Germline transformation of Aedes fluviatilis (Diptera:Culicidae) with the piggyBac transposable element Flávia Guimarães Rodrigues, Sabrina Barbosa Oliveira, Bruno Coelho Rocha, Luciano Andrade Moreira+ Laboratório de Malária, Centro de Pesquisas René Rachou-Fiocruz, Av. Augusto de Lima 1715, 30190-002 Belo Horizonte, MG, Brasil +Corresponding author: luciano@cpqrr.fiocruz.br Financial support: TDR/WHO (grant A20741) FG Rodrigues, BC Rocha, and LA Moreira: CNPq research scholarships; SB Oliveira: Fapemig research scholarship Received 25 July
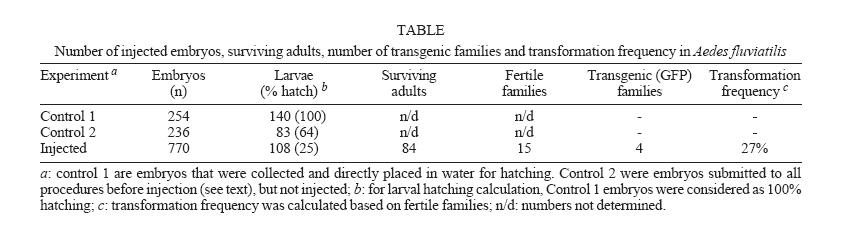
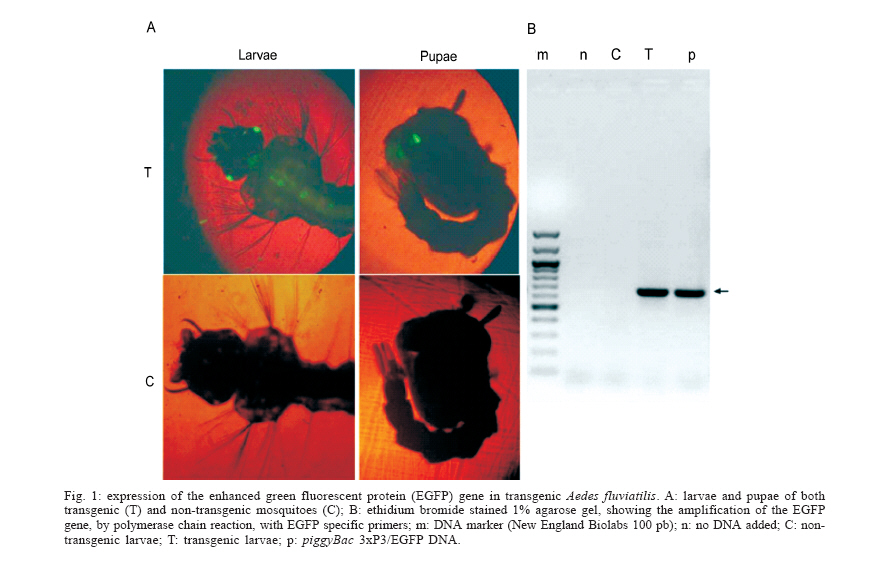
2006 Code Number: oc06126 The technique to generate transgenic mosquitoes requires adaptation for each target species because of aspects related to species biology, sensitivity to manipulation and rearing conditions. Here we tested different parameters on the microinjection procedure in order to obtain a transgenic Neotropical mosquito species. By using a transposon-based strategy we were able to successfully transform Aedes fluviatilis (Lutz), which can be used as an avian malaria model. These results demonstrate the usefulness of the piggyBac transposable element as a transformation vector for Neotropical mosquito species and opens up new research frontiers for South American mosquito vectors. Key words: transgenic mosquitoes - piggyBac - malaria The insertion of foreign genes into the genome of an organism is an important tool to address the expression of a particular gene, to study parasite-vector interactions and also for practical uses. The first stable transformation of a mosquito was achieved by Coates et al. (1998) and by Jasinskiene et al. (1998) who expressed a species-specific eye colour gene, in Aedes aegypti (Linnaeus). A great achievement was obtained with the use of the green fluorescent protein (GFP), as a selective marker, because it has the advantage to express in a variety of species, including insects (Pinkerton et al. 2000, Horn & Wimmer 2000, Catteruccia et al. 2000, Kokoza et al. 2001), and on early developmental stages. Furthermore, with the discovery that the transposon piggyBac was able to work in different organisms [insects (Handler & Harrell 1999, Handler & MacCombs 2000, Grossman et al. 2001, Peloquin et al. 2000, Tamura et al. 2000, Ito et al. 2002, Moreira et al. 2002, Kim et al. 2004, Franz et al. 2006) and even in mouse (Ding et al. 2005), planaria (Gonzalez-Estevez et al. 2003), and Plasmodium falciparum (Balu et al. 2005)], a wide horizon has opened on the transformation of insects of both agricultural and medical/veterinary importance. Although several mosquito species have been transformed so far, there are differences related to each species and consequently, adaptation of the transformation technique is required. Aedes fluviatilis (Lutz) is a zoophilic and anthropophilic species, with a geographical distribution comprehending the Southern part of Mexico to the Northern part of Argentina, east of Andes. This species has being used in laboratory as a safe avian malaria vector (Tason de Camargo & Krettli 1978), as it does not naturally transmit dengue or yellow fever. Here we report the first successful stable transformation of Ae. fluviatilis, by using the piggyBac transposable element, being the first report of genetically manipulation of mosquitoes in Latin America. MATERIALS AND METHODS Ae. fluviatilis mosquitoes were reared at 27°C and 80% humidity under a 12 h light/dark cycle. For adults, 10% sucrose solution was offered ad libitum and females were fed on mouse blood. Larvae were fed on fish food (Goldfish Colour, Alcon). The transformation plasmids [piggyBac 3xP3-EGFP (Horn & Wimmer 2000) and phsp-Helper plasmid] were purified by using a Plasmid Maxi-Prep kit (Qiagen). Plasmids were mixed to a final concentration of 0.3 µg/µl (piggyBac) and 0.2 µg/µl (Helper plasmid) in injection buffer (5 mM KCl, 0.1 mM Na2HPO4, pH 6.8) plus 5% (vol/vol) of green food colour to help visualization of injection mixture. Three days old females were fed on mouse blood and two days after, eggs were collected by forced egg laying. Thirty to 45 min embryos were lined up with their posterior ends towards the same side onto a square of filter paper soaked with isotonic buffer (150 mM NaCl, 5 mM KCl, 10 mM HEPES, 2.5 mM CaCl2, pH 7.2), the filter paper was dried-up and the embryos were transferred to a glass slide containing double-sided tape and covered with halocarbon oil 27 (Sigma-Aldrich). Microinjections were performed by using a Femtojet injector (Eppendorf) and an inverted microscope (TS100, Nikon) attached to a micromanipulator (Narishige). Microinjection parameters were: 600 hPa (injection pressure); 100 hPa (back pressure) and manual control of injection time. Quartz capillaries were pulled on a laser-based puller (Sutter) using the folllowing pulling parameters: Heat = 750, Fil = 4, Vel = 40, Del = 125, Pull = 125 and the tip was broken by touching the first embryo to be injected. Surviving injected larvae were maintained in filtered tap water until adulthood. Injected males were mated with colony-originated females and injected females with colony counterparts forming injected G0 families. Four egg batches were collected from each family and the larvae screened on a FITC-fluorescence microscope (Nikon TS-100F). Transgenic lines expressing the EGFP marker were crossed with colony insects for expansion. EGFP specific oligonucleotides (EGFPfor: 5'-ggtgcccatcctgg tcgagc-3' and EGFPrev: 5'-gcggtcacgaactccagcagg- 3') were used to identify the presence of the transgene with the following polymerase chain reaction (PCR) (95°C 2 min, 30 cycles of 95°C 1 min, 63°C 1 min and 72°C 1 min, followed by 72°C, 5 min). RESULTS AND DISCUSSION In order to adapt the microinjection technique, which has been used to other Culicidae species we tested different parameters to obtain the best larval hatching after the microinjection procedure and consequently, to increase the chances of transformation. First, we have tested the possibility of maintaining the embryos on ice to reduce embryo aging and consequently to expand the injection time, but that procedure drastically reduced larval hatching even in control embryos, which were not injected (data not shown). We tested post-injection manipulations of mosquito embryos, in order to obtain higher larval hatching. We compared the transference of embryos to a damp filter paper with the use of a fine paintbrush prior to transfer the eggs to water or by dipping the slide containing eggs in ddH20 three times, to remove oil excess. The highest larval hatching was obtained when the slide containing eggs was dipped in water to remove oil excess and the embryos were transferred to a Petri dish filled with water and with filter paper on its border. With the 84 adults that survived microinjection, 15 G0 families were formed and after screening of more than 17,000 larvae (G1), four transgenic lines were obtained (Table), resulting in a transformation frequency of 27%, based on G0 families. Fig. 1A shows the typical phenotypic characteristic of transformed mosquitoes with the use of 3xP3 promoter (Horn & Wimmer 2000) when driving the expression of EGFP. Larval, pupal and adult eyes, larval neural tube and larvae anal papillae exhibits a strong green fluorescence [similar to previously shown for Ae. aegypti (Kokoza et al. 2001) and for Anopheles stephensi (Liston) (Ito et al. 2002)], which is detectable even on first instar larvae. The presence of the EGFP gene in transgenic larvae is shown on Fig. 1B, by PCR, in comparison to non-transformed individuals. This work shows the functionality of the piggyBac transposable element, as a transformation vector, in another mosquito species. Our findings open new possibilities to genetic manipulate South American mosquito species to better understand their interaction with parasites they transmit, in our particular conditions. Work is in progress in adapting this technology to transform a Brazilian malaria vector, Anopheles aquasalis (Curry), which shall express a Plasmodium-blocking molecule. ACKNOWLEDGEMENTS To Dr Marcelo Jacobs-Lorena, who kindly provided us the transformation plasmids. To Maira S Neves and Geraldo F Carvalho for their great technical support and Cristiana FA Brito for critical reading. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06126t1.jpg] [oc06126f1.jpg] |
| |||||||||

{kind=link}
{kind=link}