 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101, No. 7, November ,2006, pp. 759-766 Genetic variability of hepatitis A virus strain HAF-203 isolated in Brazil and expression of the VP1 gene in Escherichia coli Marcia L Baptista/+, Messias Silva, Maria Amélia de Lima, Clara FT Yoshida, Ana Maria C Gaspar*, Ricardo Galler** Laboratório
de Hepatites Virais +Corresponding author: marcialb@ioc.fiocruz.br Received 26 April
2006 Code Number: oc06127 The hepatitis A virus (HAV) HAF-203 strain was isolated from an acute case of HAV infection. The primary isolation of HAF-203 in Brazil and its adaptation to the FRhK-4 cell lineage allowed the production of large amounts of viral particles enabling molecular characterization of the first HAV isolate in Brazil. The aim of our study was to determine the nucleotide sequence of the HAF-203 strain genome, compare it to other HAV genomes and highlight its genetic variability. The complete nucleotide sequence of the HAF-203 strain (7472 nucleotides) was compared to those obtained earlier by others for other HAV isolates. These analyses revealed 19 HAF-specific nucleotide sequence differences with 10 amino acid substitutions. Most of the non-conservative changes were located at VP1, 2C, and 3D genes, but the 3B region was the most variable. The availability of HAF-203 complementary DNA was useful for the production of the recombinant VP1 protein, which is a major determinant of viral infectivity. This recombinant protein was shown by enzyme-linked immunoassay and blotting, to be immunogenic and resemble the native protein, therefore suggesting its value as a reagent for incorporation into diagnostic tests. Key words: hepatitis A virus (HAV) - HAF-203 strain - complete genome (Brazil) - VP1 gene recombinant protein - Escherichia coli expression - HAV diagnosis The hepatitis A virus (HAV) is the causative agent of an old and common human disease, with high incidence rates, still present in many countries throughout the world. In Brazil, this disease has been responsible for 50% of the acute hepatitis cases referred to the National Reference Center for Viral Hepatitis/Oswaldo Cruz Institute/Ministry of Health. The etiological agent was first identified by Feinstone et al. (1973), and the virus is currently classified as the first and unique member of the genus Hepa-tovirus, Picornaviridae family (Minor 1991). HAV is a 27-nm
non-enveloped icosahedral particle, which comprises a single stranded plus-sense
RNA (Coulepis et al. 1981). The viral genome is composed of approximately
7500 nucleotides, edged by a covalently linked VPg protein at the 5' terminal
and a polyadenylated 3' end. The viral RNA encodes a single polyprotein of
2224 amino acids, which is later cleaved into 11 viral polypeptides (Cohen
et al. 198 The Brazilian HAF-203 strain investigated here was obtained from a faecal specimen of an acute hepatitis A patient. The viral isolation and serial passage history was previous reported by Gaspar et al. (1992). The HAF-203 HAV strain was inoculated into fetal Rhesus kidney cells (FRhK-4) and then reinoculated into new cultures on the 28th day and subsequently every 35 days, using tissue culture fluid (TCF) as inoculum. An HAV antigen could be demonstrated in TCF 49 days after inoculation, applying enzyme immunoassays (EIA), immunofluorescence (IF), and hybridisation tests. In addition, HAV replication was demonstrated by the detection of viral replicative intermediate (minus strand), employing reverse transcription followed by polymerase chain reaction (RT-PCR) (Baptista ML et al. unpublished). In the present study, we have sequenced the complete genome of the HAF-203 HAV strain and compared it with other HAV isolates. Several groups have adopted recombinant viral proteins for diagnostic purposes (Ostermayer et al. 1987, Johnston et al. 1988, LaBrecque et al. 1998, Ternovai et al. 2001, Di Napoli et al. 2004). A fragment of the HAF-203 strain encoding the complete VP1 gene was cloned and expressed in Escherichia coli, to provide valuable specific immune reagents for both diagnostic and epidemiological surveys of HAV infection. MATERIALS AND METHODS Virus -
Virus from the 4th passage in FrhK-4 cells was purified from culture medium
and infected cells. Culture medium was collected weekly, during 28 days,
and stored at _20°C. Monolayer infected cells (28th passage after inoculation)
were suspended in lysis buffer (10 mM Tris buffer pH 7.5 containing 50 mM
NaCl, 0.5% NP40, 0.5% deoxycholic acid, and sodium salt - DOC) then frozen
and thawed three times. The cellular extracts were centrifuged at 6000 g
for 10 min and re-extracted twice with lysis buffer. Culture medium and cellular
extracts were combined and precipitated, using 0.3 M NaCl _ 10% polyethylene
glycol 6000. The mixture was stirred at 4°C overnight, and the precipitate
was collected by centrifugation at 10,000 g for 30 min. The supernatant was
discarded, and the precipitate was suspended in 10 mM Tris buffer (pH 7.5),
containing 50 mM NaCl and 1 mM EDTA (TNE buffer). A new extraction was carried
out using trichlorotrifluoroethane (FREON), and the aqueous phase was separated
by centrifugation at 5000 g for 10 min. The aqueous phase was reserved, and
the FREON phase and interface were re- RNA extraction - Sodium dodecyl sulphate (SDS) and b-mercaptoethanol in a final concentration of 1% were added to every aliquot of HAF-203 strain concentrated in sucrose cushion. RNA extraction was performed applying phenol-chloroform followed by ethanol precipitation. cDNA synthesis
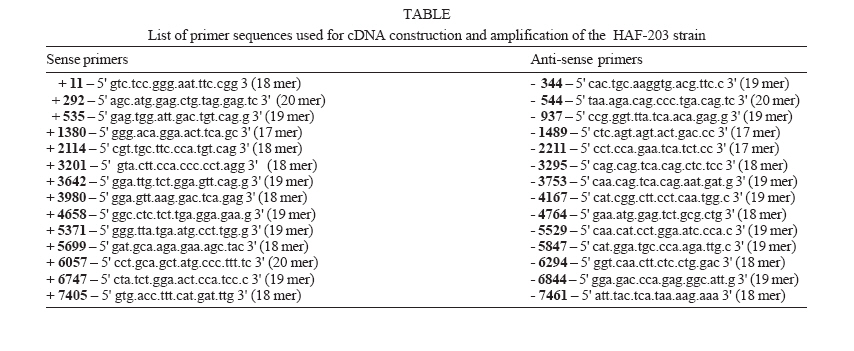
(reverse transcription) and PCR 1 - A total of 28 specific primers
(Table) were employed for reverse transcription
and cDNA amplification by PCR. For complementary DNA (cDNA) synthesis,
RNA was res-suspended in water and mixed with negative-sense primer (0.1
pmol), corresponding to HAV nucleotide sequences. After heating at 90°C
for 2 min and slow cooling down to room temperature (5 min), reverse transcription
was carried out for 60 min at 37°C, using the reaction mixture: 50
mM Tris-HCl pH 7.3, 50 mM KCl, 10 mM MgCl2, 5 mM DTT, 0.2 mM
each deoxynucleotide, 400 U/ml Rnasin (BRL), and 200 U/ml MMLV PCR 2 - The first PCR products were isolated by electrophorectic separation on low-melting temperature agarose (Sigma), and a second PCR was carried out following the same protocol, except for the number of cycles (25 cycles). Cloning - The PCR 2 products were cloned into a pGEM 2 (Promega) or pCRII (Invitrogene) plasmids. For the recombinant protein expression, a fragment carrying the VP1 gene was cloned into a pET28a plasmid (No-vagen). Sequencing - Nucleotide sequencing was performed according to the dideoxy termination method (Sanger et al. 1977) using SP6 and T7 primers (Promega). Sequenase (USB) and [32P] dATP (ICM) were utilized, according to manufacturer's specifications. Sequencing reactions were submitted to electrophoresis in 0.4 mm-thick Tris-Borate-Urea gels and visualized by autoradiography. At least two clones of each region were sequenced on both strands consisting of 12 clones and 13 sub-clones along the 7500 nucleotides of the HAF-203 genome. When differences were observed, additional clones were evaluated. Nucleotide sequence analysis - For this purpose, we selected one complete nucleotide sequence from each HAV (genotype I) including the HM-175 strain from Australia (Cohen et al. 1987b) GenBank accession number M14707, to which the nucleotide sequence of its attenuated derivative (Cohen et al. 1987a), accession M16632, was also included. The other sequences, available in databanks, were: the LA strain from Los Angels/US (Najarian et al. 1985) accession K02990; the MBB strain from Africa (Paul et al. 1987) accession M20273; the HAS-15 strain from US (Sverlov et al. 1987) accession X15464 and X15463; the GBM strain from Germany (Graft et al. 1994) accession X75215; the HAV-FG strain from Italy (Beneduce et al. 1995) accession X83302. All analyzed strains were adapted to cell cultures. The HAF-203 nucleotide GenBank accession number is AF268396. Sequence analysis was performed, adopting the "Seqed and Fasta" programs (Wisconsin University, Sequence Analysis Package, GCG, Madison, WI), and Clustal X software (Thompson et al. 1977). Phylogenetic tree - The tree was generated using the Kimura two-parameter model with full-length genomic sequences and neighbor-joining method, MEGA 3.1 version (Kumar et al. 2004). The reliability was assessed by bootstrap re-sampling (1000 pseudo-replicas), also in MEGA. The nucleotide sequences from the following strains were retrieved from GenBank: AGM 27 (D00924), AH1 (AB020564), AH2 ( AB020565), AH3 (AB020566), CF53 (AY644676), DL3 (AF512536), FG (X83302), FH1 (AB020567), FH2 (AB020568), FH3 (AB020569), GBM (X75215, X75214, and X75216), HAF-203 (AF 268396 this paper), HM-175 (M14707, NC001489), HM-175 attenuated (M16632), HAS-15 (X15464 and X15463), IVA (DQ646426), LA (K02990), LY6 (AF485328), M2 (AY974170), MBB (M20273), NOR-21 (AJ299464), and SLF88 (AY644670). Bacterial expression -
The DNAs encoding the complete VP1 (the major viral antigen), parts of 2A
and VP3 HAV proteins were cloned and expressed in E. coli. A 1.2 Kb
fragment (amino acid 461-860) was obtained by enzymatic digestion from the
42pGEMVP1 clone. After digestion with Eco RI and Hind III (restriction sites
present in plasmid pGEM2), the fragment was inserted into the expression
vector pET28a (Novagen). The resulting plasmid was referred to as VP1pET42u.
In addition, a vector control of the expression procedure, plasmid VP1pET85t,
was employed. It constituted another pET construct with the same fragment,
not in frame. For bacterial expression of the recombinant protein, E.
coli strain BL-21 (DE3) was transformed using the pET28a constructs.
The cultures were incubated at 37°C in Luria broth, containing kanamicin
(50 µg/ml) until OD600 = 0.8. For the induction step, we
added isopropyl-b-D-thiogalactopyranoside (IPTG) to a final concentration Purification of VP1pET42u and measurement of concentration - The His-tagged recombinant protein was purified by immobilized metal-ion affinity chromatography (IMAC) using six histidine residues added to the carboxi-terminus of the VP1 recombinant protein. Briefly, the His-tagged VP1PET42u was extracted from the bacterial pellet under denaturing condition (6M urea), bound to Ni2+ (Ni-NTA superflow - Qiagen) and eluted with 100 mM imidazole, following the manufacturer's instructions (The QIAexpressionist Kit - Qiagen). The purified protein solution was quantified by the Lowry method (Lowry et al. 1951), standardized with bovine serum albumin and estimated by molar extinction coefficient at 280 nm. Rabbit immunization - Rabbits were immunized with the recombinant VP1 protein. The protocol for immunization was adopted as previously described by Camargo et al. (1987). Immune serum was obtained after four 80 µg intramuscular doses of purified recombinant VP1 protein at two week intervals. Conjugation of anti-HAV rabbit immune serum to horseradish peroxidase - The resulting antiserum was assayed by EIA for the detection of anti-HAV antibodies (Hepanostika HAV Antibody Biomerieux). Rabbit anti-HAV gammaglobulin was precipitated with ammoniun sulphate, according to Hebert et al. (1973). Four milligrams of purified IgG was conjugated to 10 mg/ml horseradish peroxidase (type IV; Sigma). The Nakane's method was adopted as modified by Carmargo et al. (1987). Dot Blot - The
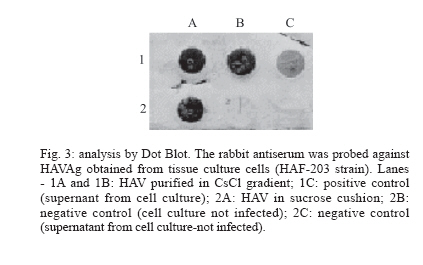
rabbit HAV antiserum was also analyzed by Dot Blot. Aliquots of 100 µl
FRhK-4 cells infected or not with HAF-203 virus were incubated (1 h at 65°C)
with 100 µl of Triton-X100-SDS buffer (50 mM Tris-HCl; 0.1% Triton-X100
and 2% SDS) extracted with phenol chloroform (1:1, v/v) and once more with
chloroform (1:1, v/v). Positive and negative feces for HAV antigen were utilized
for testing the HAV antiserum. Stool samples were prepared as a 10% (w/v)
suspension in PBS pH 7.2, clarified by low-speed centrifugation and treated
as described above. Samples were boiled at 100°C for 5 min and applied
directly to nitrocellulose paper (BioRad) using a Minifold II slot-blot apparatus.
After heating for 2 h at 65°C in a vacuum oven, the membrane was incubated
in PBST-Milk solution (phosphate buffer - PBS pH 7.2; 0.05% tween 20 plus
5% dried non fat milk) for 16 h at room temperature. The membrane was washed
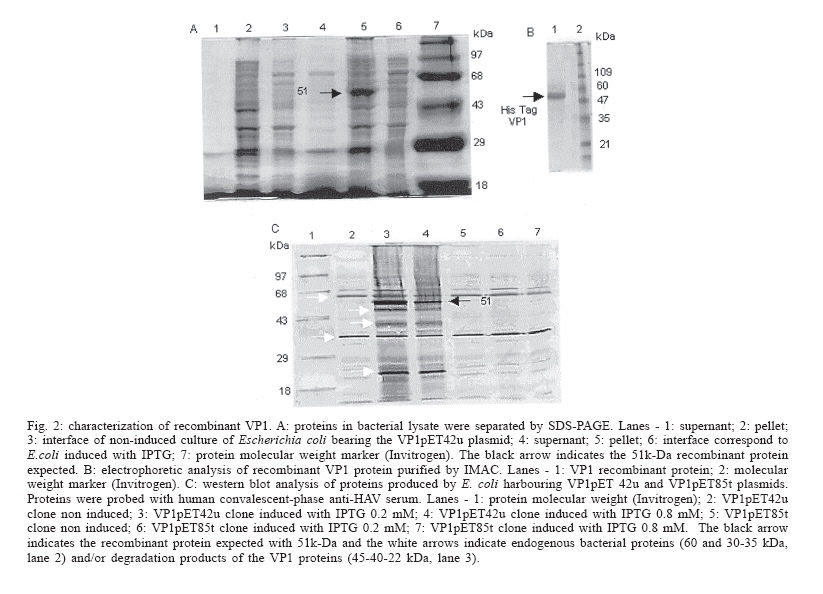
in 15-20 ml Competitive Immunoassay test with rabbit anti-recombinant VP1 horseradish peroxidase conjugate - A standard protocol for the detection of antibodies against HAV was adopted (Vitral et al. 1991) to analyze the rabbit anti-VP1 conjugate. Briefly, a gammaglobulin IgG anti-HAV was used to coat a polystyrene well plate exposed to overnight incubation at 8°C. Further, an HAF-203 antigen from culture supernatants and positive control sera (anti-HAV IgG) were simultaneously added. After incubation and washing steps addition of rabbit anti-VP1-horseradish peroxidase conjugate was succeeded. Tetramethylbenzi-dine (TMB) plus H2O2 were employed to reveal the reaction followed by spectrophotometric reading at 450 nm. The cut-off value was defined as the mean of positive and negative control values. Specimens with absorbance values greater than the cut-off value were considered as negative and those with absorbance values lower than or equal to the cut-off were considered as a positive result. RESULTS Nucleotide and amino acid sequence heterogeneity in the Brazilian HAF-203 strain genome - The 7472 nucleotides of the HAF-203 strain genome were sequenced in both strands and at least two clones for each region were investigated. In the HAF-203 strain, the nucleotide changes were distributed throughout the genome and most of them constituted silent mutations with VP1, 2C, and 3D having accumulated most of the nucleotide and amino acid changes. Nineteen nucleotide
alterations, exclusive to the HAF-203 strain, were present along the genome,
however only 10 of them resulted in amino acid substitutions. Mutations were
observed in the VP3 protein (nucleotide 1574: Asp→ Val); VP1 protein
(nucleotide 2292: Ile→Met, nucleotide 2941: Glu→Lys and nucleotide
3031: Val→Pro); 2A protein (nucleotide 3294: Ile→Met); 2B protein
(nucleotide 3724: Asp→His and nucleotide 3808: Phe→Leu); 3B protein
(nucleotide 5268: Gln→His) and 3D protein (nucleotide 6164: Asp or Asn→Gly
and nucleotide 7093: Thr→Ala). In addition, three other non-exclusive
nucleotide changes which led to amino acid substitutions were observed in
the 2C protein (nucleotide 4439: Lys→Thr and nucleotide 4871: Asn→Ser)
and in the polymerase gene (3D) (nucleotide 6555: Ile→Met). Six silent
nucleotide changes were present in the VP3 protein (nucleotide 1752 shifted
from G→A), in theVP1 protein (nucleotides 2703 and 2871 mutated from
T→A Genetic relationship between HAF-203 and other HAV strains - Our results have demonstrated that the HAF-203 strain belongs to genotype IB (Fig. 1). In relation to the genetic diversity, our data suggest that HAF 203 strain presents a high degree of homology with the HM-175 (accession - M14707) and HM-175 attenuated (accession - M16632) strains, with 99.7 and 99.5% of identity, respectively. Furthermore, an identity of 94.8% presented itself between HAF 203 and MBB (accession - M20273) strains with about 91% of homology. When the remaining strains were considered (CF53/Berne - accession AY644676; SLF88 accession - AY644670; NOR-21 - accession AJ299464, and AGM-27 strains accession - D00924), a similarity of 86.3, 86, 83, and 81.2% was observed. Expression of VP1 in E. coli - We selected the VP1 protein for expression in E. coli since it is the main viral capsid protein from structural and immunogenic standpoints. The recombinant plasmids containing the entire VP1 gene from the HAF-203 strain produced in E. coli was analyzed by SDS-PAGE and Western blot. The expression of a 51 kilodalton (kDa) recombinant protein was observed in bacterial extracts previously transformed with the VP1pET42u plasmid (Fig. 2A). However, the expression of this protein was not detected in bacteria containing the VP1pET85t clone, a plasmid with HAF-203 VP1 gene cDNA inserted in the opposite orientation. The yield of recombinant VP1was determined to be approximately 2 mg/l of bacterial culture (Fig. 2B). In order to investigate the antigenicity of the recombinant VP1, its reactivity with anti-HAV serum was valuated by Western blot (Fig. 2C). The recombinant 51-kDa protein was specifically recognized by human convalescent antiserum to HAV. Rabbit HAV antiserum - There was a seroconversion of two rabbits immunized with recombinant VP1 protein was analyzed with the Hepanostika HAV Antibody assay (Biomérieux). The OD values of the positive controls ranged from 0.184 to 0.209 whereas those of the negative controls ranged from 0.759 to 0.828. Anti-HAV sera OD values from the two immunized rabbits were 0.121 and 0.139, respectively, (cut-off value of 0.495). According to dot blot analysis the rabbit HAV antiserum reacted specifically with the HAVAg obtained from tissue culture cells (Fig. 3). DISCUSSION We have sequenced the complete genome of the HAV HAF 203 strain, previously isolated in a cell culture system (Gaspar et al. 1992) in order to assess its genetic variability as compared to other known strains. Mutations were observed along the HAF 203 genome corroborating previous reports in which the authors found mutations scattered throughout the viral genome of HAV strains adapted to cell culture (Najarian et al. 1985, Cohen et al. 1987a,b, 1989, Paul et al. 1987, Graft et al. 1994, 1997, Beneduce et al. 1995). A higher frequency of mutations occurred in the 3B region, ranging from 4.3 to 8.6%. The VP1, 2C, and 3D protein genes of HAF-203 displayed non conservative amino acid substitutions, which may have adaptation value for the HAF-203 virus to replicate in cell cultures, but they did not affect its infectivity to non human primates (Vitral et al.1995, Pinto et al.2002). Ten nucleotide
changes led to amino acid substitutions, which are unique in the HAF-203
strain. Those at residues 2292 (VP1), 3294 (2A), and 3808 (2B) led to conservative
amino acid substitutions and are not likely to have markedly altered these
proteins. However, one nucleotide substitution in the VP3 gene (nucleotide
1574), two in VP1 (at positions 2941 and 3031), one in 2B gene (nucleotide
3724), one in 3B protein (at nucleotide 5268) and two in the 3D protein gene
(nucleotides 6164 and 7093) may be relevant to cell culture adaptation. It
was also observed that two nucleotide changes in the 2C protein from the
HAF-203 strain (at nucleotides 4439 and at 4871) exhibited the same amino
acid substitution as in the HAV-FG strain. The mutation at nucleotide 4872
(Asn→ Met) was correlated to HAV propagation in cell culture (Beneduce
et al. 1995). On the other hand The occurrence of deletions and insertions has been reported during the alignment of full-length HAV genomic sequences. Deletions were observed in the 5'NC region and in VP1 and 3A proteins, while insertions were observed in the 5'NC region of the VP1 protein. Beneduce et al. (1995) and Graft et al. (1997) suggested that these findings may be important when isolated HAV is being adapted to cell cultures. It will be of interest to compare the molecular basis of HAF-203 strain adaptation to FrhK-4 and to a new cellular substrate (certified Vero cells). This virus is now being adapted for growth in Vero cells, with an aim toward the development of an inactivated vaccine against hepatitis A. A previous report has suggested the existence of at least seven distinct genotypes for HAV (Robertson et al. 1992. In an attempt to better characterize the HAF-203 genotype, its nucleotide sequence was compared to 25 complete nucleotide sequences available in databanks. The HAF-203 strain displays a high degree of homology with HM-175, its attenuated derivative, IVA and MBB strains (99.7, 99.5, 98.8, and 94.8% respectively), confirming previous data (de Paula et al. 2002, Lu et al. 2004, Stene-Johansen et al. 2005) in which HAF-203 was assigned to genotype IB. In relation to other isolates (GBM, HAV-FG, M2, HAS-15, LA, FH1, AH2, AH3, LY6, FH3, DL3, AH1, and FH2 strains) the level of homology found in comparison with the nucleotide sequence of the HAF-203 strain varied from 91.6 to 90.7%. These strains are classified as genotype IA and they are the most genetically divergent from the HAF-203 strain. When our isolate was compared with CF53/Berne and SLF88, a nucleotide homology of 86.3 and 86%, respectively, was resulted. These strains belong to genotype IIA and IIB, respectively. The level of homology encountered between the NOR-21 and HAF-203 strains in the nucleotide sequence was 83%, NOR-21 being classified as genotype IIIA. Finally, when our isolate was compared with the AGM-27 strain, a nucleotide homology of 81.2% was observed. These results confirm previous studies in which the AGM-27 strain proved to be unique, because it differs significantly from any other HAV isolate (Lemon et al. 1987, 1992, Tsarev et al. 1991), such as the CF53/Berne, SLF88 and NOR-21 isolates (Ching et al. 2002, Lu et al. 2004, Stene-Johasen et al. 2005). Our results confirm
previous studies where strains from the southern hemisphere, such as HAF-203,
HM-175 and MBB, have a very close nucleotide identity while strains from
the northern hemisphere are more related to each other. The HAF-203 strain
as well as the HM-175 and MBB strains, more than likely, have a common geographic
origin. For the expression
of the HAF-203 VP1 protein in E. coli two recombinant plasmids carrying
the VP1 gene were constructed. Our results revealed that human serum recognized
the 51k-Da recombinant protein present in E. coli extracts in contrast
with a previous report (Ostermayer et al. 1987), which claimed that VP1 in E.
coli was not antigenic. Two bands of approximately 60 and 30-35 kDa probably
reflect recognition of endogenous bacterial proteins by the human serum.
The other bands (45-40-22 kDa) might correspond to degradation products of
the VP1 protein expressed in E. coli as suggested by Johnston et al. Previous studies have demonstrated that either VP1 (Stapteton & Lemon 1987) or VP3 (Ping et al. 1988) proteins contain the most important neutralization epitopes in the HAV virion. Recombinant HAV VP1 and VP3 expressed in E. coli have also proven to be antigenic in rabbits (Johnston et al. 1988, Gauss-Müller et al. 1990, Powdrill et al. 1991). We have immunized rabbits with the recombinant VP1 protein and the resulting antiserum was conjugated to peroxidase. Our results indicate that the anti HAV rabbit sera peroxidase conjugate may be applied, after appropriate standardization, as a reagent in a competitive enzyme immunoassay for HAV infection diagnosis. ACKNOWLEDGMENT To Dr José Antônio Pinto Sá Ferreira (Diagnosis Reactive Department of Bio-Manguinhos-Fiocruz) for his help in reviewing the manuscript. Reviewed and revised by Mitchell Raymond Lishon, native of Chicago, Illinois, US, UCLA 1969. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06127f1.jpg] [oc06127t1.jpg] [oc06127f3.jpg] [oc06127f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}