 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101, No. 7, November ,2006, pp. 795-799 Synonymy between Lymnaea bogotensis Pilsbry, 1935 and Lymnaea cousini Jousseaume, 1887 (Gastropoda: Lymnaeidae) Luz Elena Velásquez Programa de Estudio
y Control de Enfermedades Tropicales, Escuela de Bacteriología y Laboratorio
Clínico, Universidad de Antioquia, Sede de Investigación Universitaria,
Calle 62 # 52-59, 7 piso, 730, Medellín, Colombia Financial support: Colciencias, grant 1115-05-13664 Received 19 June
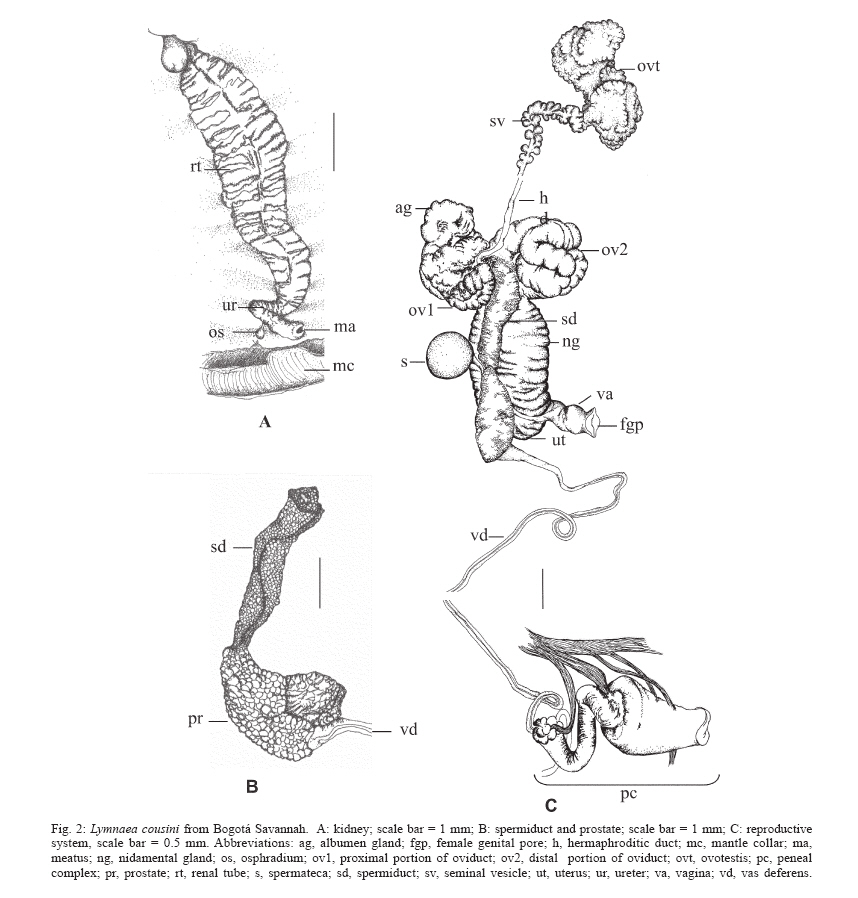
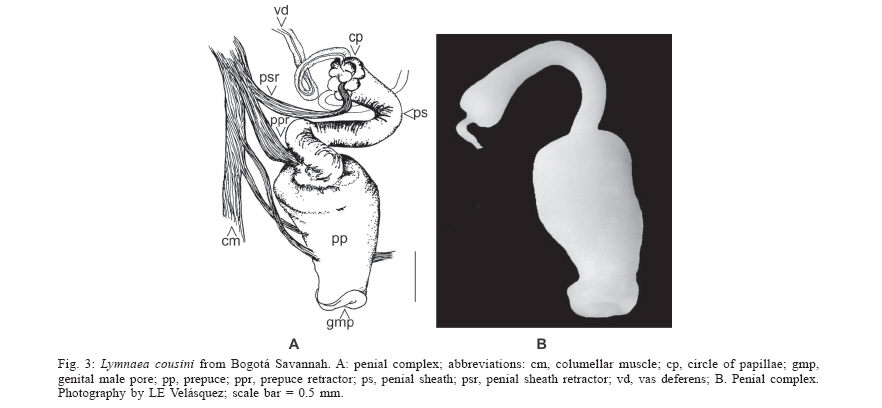
2006 Code Number: oc06133 The lymneid snail Lymnaea bogotensis Pilsbry 1935 is synonymized with L. cousini Jousseaume 1887, based on morphological comparisons of the reproductive systems. The shell, renal tube, and reproductive system are described and illustrated from specimens collected in the type locality and the municipality of Paipa, Colombia. Bibliographical records reveal L. columella to be the commonest lymneid in the country. The latter two species can be differentiated based on the morphology of the penial complex. Key words: Lymnaea bogotensis - Lymnaea cousini - synonymy - taxonomy - Colombia Members of the family Lymnaeidae are pulmonate molluscs present in continental aquatic ecosystems throughout the world. About 100 species have been described, most in the Holarctic region with a few representatives in the Neotropics (Hubendick 1951). A few species of lymneids are important in medical malacology because of their roles in the transmission of digeneid parasites such as Fasciola spp. and Paramphistomum spp., responsible for fasciolosis in bovines and humans as well as paramphistomosis in ruminants (Malek 1985). Since these diseases generate huge economic losses in livestock production in tropical and subtropical countries of all continents, intervention methods are needed to control their expansion and reduce their respective prevalences (WHO 1995). Populations of lymneid species that act as hosts for these parasites also become the object of epidemiological vigilance. Furthermore, studies to identify potential vectors and map their distributions are important in determining zones at risk from disease. Paraense (1976a, 1982a, b, 1983, 1984, 1995, 2004) made the most important taxonomic studies of South American Lymnaea, describing the morphological characters of the reproductive system, kidney, and shell of each species in detail. This methodology was applied by Oviedo et al. (1995) and Samadi et al. (2000) in their studies on lymneids of the Bolivian plateau, the results of which were confirmed by isoenzyme and molecular techniques (Carvalho et al. 2004). Pointier et al. (2004) also used this methodology in studies of Venezuelan species. Three species of Lymnaea have been reported in Colombia, including L. ubaquensis Piaget, 1914, and L. bogotensis Pilsbry, 1935; new species whose respective type localities are Laguna de Ubaque and Bogotá, both in the department of Cundinamarca above 2500 masl. The latter species was incriminated by Brumpt et al. (1940) and Muñoz-Rivas (1953) as host of Fasciola hepatica in this region. It has been suggested that both species are synonyms of L. cousini (Hubendick 1951, Pointier et al. 2004). On the other hand the lymneid that presents the widest geographical distribution in Colombia is L. columella (Say 1817), which inhabits aquatic ecosystems from sea level to 2600 masl and has been identified as an intermediate host of F. hepatica (Patiño 1978, Malek & Congswell 1980, Malek 1985, Gómez 1990). Despite their epidemiological importance, there is considerable confusion in the systematics of Colombian lymneids. The formal descriptions of L. ubaquensis and L. bogotensis include photographs and some characteristics of the shells but lack detailed descriptions and drawings of their internal structures, needed to help establish the status of each as a species. This paper confirms that L. bogotensis Pilsbry 1935 is synonymous with L. cousini Jousseaume 1887, based on morphological comparisons. L. bogotensis Pilsbry (1935: 88. Plate 6, Fig. 9) was briefly described as follows: "Bogotá. Type 164556 A.N.S.P., collected by Hmo. Apolinar Maria. The shell is umbilicate, ovate, thin, light brown, with short, conic spire; glossy, with fine, irregular striae and wrinkles of growth, which, above the periphery, are cut into long granules by some indistinct spirals. The large aperture is oblique, ovate, broadly rounded below. Columella dilated above, passing in a very wide angle into the thickened parietal callus. There is a distinct, long fold, chiefly visible in an oblique view in the mouth. Length 13.7 mm., diam. 8.6 mm., length aperture 9.8 mm". MATERIALS AND METHODS Type locality - Five specimens of L. bogotensis in formaldehyde (shells and soft tissues) were obtained from the collection of the Instituto Nacional de Ciencias of the Universidad Nacional de Colombia. These snails were collected in 1974 in the south of Bogotá (4º 35' 56" N; 74º 04' 51" W). Other locality - Snails were collected from a stream, the Quebrada Toibita (5º 47' 04" N; 73º 06' 47" W) at Paipa (Boyacá) in May and December 2003. They were transported live to the laboratory and maintained on fresh lettuce in aquaria provided with artificial aeration, dechlorinated water and sterile mud substrate. Thirty adult specimens were selected for taxonomic studies. The snails were relaxed in 0.05% Nembutal for 8 h and sacrificed by immersion in water at 70ºC. The soft tissues were separated, fixed in Railliet-Henry solution and dissected following the methodology proposed by Deslandes (1951) and Pa-raense (1966, 1975, 1976b, 1988). Measurements of the reproductive system were carried out with a micrometer linked to an optical microscope (± 0.01 µm). Shell measurements were taken with a digital calibrator (± 0.01 µm) and stored in recipients sealed with cotton wool. The kidney and reproductive system of each specimen were illustrated using a camera lucida. Specimens from the type locality and Paipa were deposited in the collection of the Programa de Estudio y Control de Enfermedades Tropicales within the Instituto Alexander von Humboldt, VHET 37, Colombia (PECET MOL 85-86, 317-319). DESCRIPTION The largest shell (Fig. 1) was 11.7 mm high and 7 mm wide; height of spire 4.6 mm, aperture 7.1 mm high and 5.3mm wide. The following ratios were calculated for the 35 specimens, with spire heights of 3.1-11.7 mm (6.85 ± 2.3): shell width/shell height = 0.54-0.71 (0.62 ± 0.05); spire height/shell height = 0.24-0.45 (0.36 ± 0.05); aperture height/shell height = 0.55-0.76 (0.64 ± 0.05); aperture height/spire height = 1.23-3.17 (1.84 ± 0.41). From these ratios it was inferred that the aperture constitutes about 2/3 of the total shell height. Qualitative characters of the shells are as follows: light brown, umbilicate, thin, with short spire and fine growth lines. Aperture large, oblique, oval and wide at the base, with a fold in the middle part of the parietal wall, on which a thin white callus can also be observed. The renal tube (Fig. 2a) extends diagonally across the roof of the palial cavity, from the pericardial region to the mantle collar, where it folds over to the left and returns as a ureter that opens out into a meatus to the right of the osphradium. The ovotestis (Fig. 2c) is a mass of compact sigmoid tubes that opens into a ovispermiduct, in which can be observed very short lateral, digitiform structures that comprise the seminal vesicle along the first two-thirds of its length. The last third of the ovispermiduct is very thin and simple and terminates in the carrefour. The milky white albumen gland lies on the carrefour and the first portion of the oviduct, hiding the origin of these structures. The oviduct is a transverse tubular structure, curled over at the extremities. Its region proximal to the albumen gland is smaller than the distal region, which is formed by larger, dilated tubes and continues as the nidamental gland. The nidamental gland is compact with transverse striations. At the base of this structure lies the uterus which has a softer aspect and vertical striations. A prolongation to one side forms the short, bulbous vagina, opening to the exterior through the female genital pore. The spermatheca is a spherical yellow structure with a canal which separates from the centre of the vagina and crosses diagonally between the prostate and the nida-mental gland. It reaches the roof of the palial cavity close to the pericardium. The spermiduct leaves the carrefour beside ov1 and continues along a depression in the middle of the ventral region of the nidamental gland. Its small cells give it a granular aspect. It opens into the prostate (Fig. 2b), which is wide and concave with folds, its cells being larger than those of the spermiduct. A funnel emerges from the dorsum of the distal region of the prostate, the vas deferens. This narrows until it attains its smallest diameter in the first third of its length; it then continues somewhat thicker and more sinuous to the penis which lies within the penial sheath (Fig. 3) whose proximal extremity is somewhat thicker and bears a ring of papillae. The penial sheath is cylindrical, with a length 1.5 ´ that of the prepuce (range 1.30-1.96, mean 1.5 ± 0.24). This is thicker at the point of insertion of the penial sheath and terminates in the male genital pore, whose borders are lightly thickened. DISCUSSION The qualitative characters of the shell described in this study coincide with those given by Pilsbry (1935) for L. bogotensis. Furthermore, the shell measurement ratios and internal structures such as the kidney and the reproductive system resemble those in the re-description of L. cousini Jousseaume, 1887 by Paraense (1995), in the specimens collected in Bogotá in 1974 as well as those from Paipa in 2003. Colombian specimens of L. cousini thus occur within the altitudinal range reported for this species in other Neotropical countries (Pointier et al. 2004, Paraense 2004) In summary, the specimens described unite a complex of characteristics that coincide exactly with those given for L. cousini by Paraense (1995). Thus L. bogotensis Pilsbry, 1935 is a synonym of L. cousini Jousseaume, 1887. Some of the specimens of L cousini collected in Paipa were parasitized with F. hepatica, which constitutes proof of its role as a host of the digeneid and supports the conclusions of other authors (Pointier et al. 2004, Villa-vicencio & Carvalho de Vasconcellos 2005). Although Physa acuta was collected in nine aquatic ecosystems in the Sabana de Bogotá by Velásquez and Escobar (2001), lymneids identified as L. columella were obtained from only one of these. The shell of this species is similar to that of L. cousini, given that in both the aperture is about 2/3 of the total height; but they differ in the characteristic microsculptures of the periostracum present in L. columella, which are absent in L. cousini (Pointier et al. 2005). The two species also resemble each other in the double flexing of the ureter, but can be differentiated by comparing the morphology of the penial complex. In L. cousini it is very conspicuous and consists of a wide prepuce with a penial sheath that is always 1.5 ´ longer, while in L. columella the prepuce is very slender and the penial sheath much shorter (0.26 ´). The prostate of L. cousini is large and voluminous while that of L. columella is very thin. This study illustrates the importance of comparative descriptions of the reproductive system, particularly the penial complex, in the taxonomy of Colombian lymnaeids. ACKNOWLEDGEMENTS To Dr Wladimir Lobato Paraense for suggesting this research topic and for his guidance and support. To Dr Iván Darío Vélez for his continued support of our research on Colombian snail vectors. To biologist Erika P Alarcón for collecting and transporting the specimens from Paipa. To María Carmenza Hincapie for the drawings. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:Photo images[oc06133f3.jpg] [oc06133f2.jpg] [oc06133f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}