 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101, No. 8, December ,2006, pp.851-856 Single and concomitant experimental infections by Endotrypanum spp. and Leishmania (Viannia) guyanensis (Kinetoplastida: Trypanosomatidae) in the Neotropical sand fly Lutzomyia longipalpis (Diptera: Psychodidae) André F Barbosa/+, Sandra MP Oliveira, Álvaro L Bertho*, Antonia MR Franco**, Elizabeth F Rangel Laboratório
de Transmissores de Leishmanioses, Departamento de Entomologia *Departamento
de Imunologia, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365,
21040-900 Rio de Janeiro, RJ, Brasil **Coordenação
de Pesquisas em Ciências da Saúde, Instituto Nacional
de Pesquisas da Amazônia, Manaus, AM, Brasil Financial support: Faperj Received
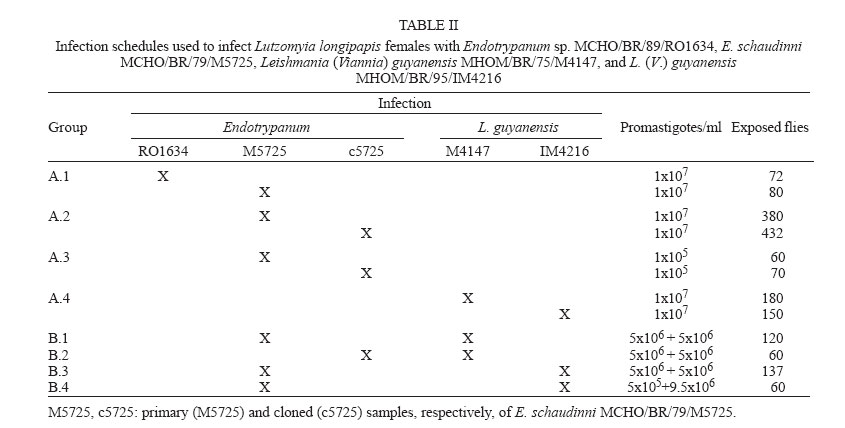
15 May 2006 Code Number: oc06140 Lutzomyia longipalpis females received single and mixed infections with Endotrypanum and Leishmania. Two biological parameters were analyzed: the percentage of infected females and the distribution of flagellates in the gut of the females. The principal comparisons were performed between (1) two strains of Endotrypanum, (2) cloned versus primary sample of one strain of Endotrypanum, (3) Endotrypanum versus Leishmania guyanensis, and (4) the pattern of flagellates behaviour by optical microscopy in females with single or mixed infection versus the identification of parasites isolated from digestive tracts by isoenzyme electrophoresis. Flagellates of Endotrypanum showed distinct patterns of infection suggesting that there is variation between and within strains. The distribution of Endotrypanum and L. guyanensis differed significantly in relation to the colonization of the stomodeal valve. In co-infection with L. guyanensis, a large number of flagellates were seen to be plentifully infecting the stomodeal valve in significantly more specimens than in females infected by Endotrypanum only. However, the electrophoretic profiles of isoenzymes of parasites recovered from all co-infected specimens corresponded to Endotrypanum. This suggests that the mere correlation sand fly infection-biochemical analysis of isolates may induce parasitological incorrect consideration. Key words: Endotrypanum - Leishmania (Viannia) spp. - Lutzomyia longipalpis - flow cytometric cloning - co-infection There are some records of concomitant infections by Leishmania and Endotrypanum in sand flies and in vertebrate hosts (Table I). This fact has considerable epidemiologic importance, mainly as some researchers consider that the morphology and the development of Leishmania (Viannia) spp. and Endotrypanum in the sand flies intestine are similar (Shaw 1992, Franco et al. 1997b). Besides, the relation between the infection observed in the sand flies intestine and the biochemical and molecular characterization of parasites can lead to a mistaken interpretation regarding the specific identification of flagellates, due to the fact that the tissue cultivation and/or host fluid originate, usually, pure isolates. According to some authors, this selection process can happen in an extremely fast way (Deane et al. 1984, Spithill et al. 1984, Ibrahim et al. 1994). Consequently, we could have been led to an incorrect estimative of natural (co)infections of sand flies by Leishmania and Endotrypanum. The objective of the present work was to compare and evaluate the behaviour and infectivity, in Lutzomyia longipalpis, of different strains of Endotrypanum and L. (V.) guyanensis, in single and mixed infections, and identify flagellates recovered in cultures by eletrophoretic profiles of isoenzymes. MATERIALS AND METHODS Sand flies - Females of L. longipalpis were collected at Lapinha Cave (19o33'40"S, 43o 57'30"W) and were reared in accordance with Rangel et al. (1986, 1987) and Wer-melinger et al. (1987). Adults were collected with CDC light traps and were shipped to Rio de Janeiro within cages. Parasites
- Endotrypanum sp. MCHO/BR/89/RO1634, E. schaudinni MCHO/BR/79/M5725,
and L. (V.) guyanensis MHOM/BR/95/IM4216 were donated by
Dr AMR Franco. Endotrypanum RO1634 and M5725 represent, respectively,
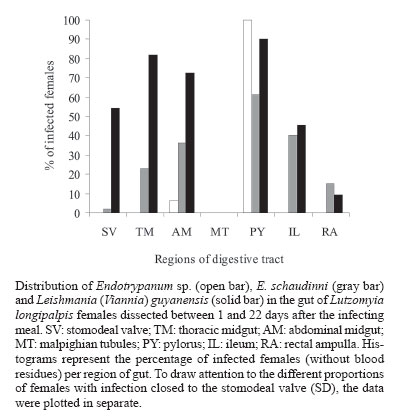
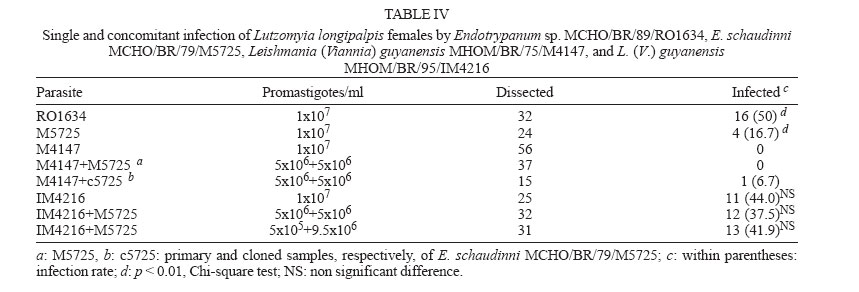
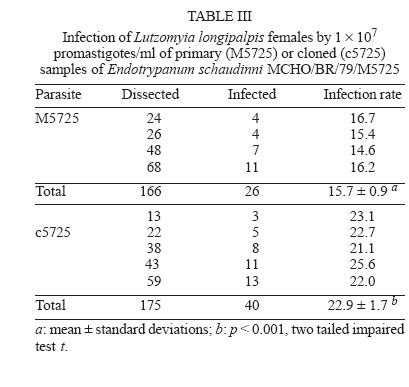
zymodeme/phenetic groups: Flow cytometric cloning - Promastigotes of E. schaudinni M5725 from exponential-phase cultures were obtained using supplemented SIM. At this time, the parasite suspension was centrifuged at 4000 rpm/10 min and washed three times in PBS, at pH 7.2. The parasites were adjusted to a concentration of 106 promastigotes/ml and were submitted for cloning using the EPICS 751 flow cytometer (Coulter, Hialeah, FL, US). An electronic gate was created, based on morphological parameters, and the procedure of single-cell sorting (cloning) was performed using the Auto-Clone device (Coulter). One parasite was placed in each well of a 96-well microtitration plate containing SIM. The plates were cultured at 26°C for seven days. The clones that grew were transferred to batch cultures to obtain parasite mass and one clone (named c5725) was selected at random. Experimental design - Female sand flies, F1 generation, 4-6 days old, randomly distributed to the experimental cages, were infected by feeding through a chick-skin membrane, obtained as programme No. P0097-01 of Fundação Oswaldo Cruz Ethics Committee on Animal Experimentation (CEUA-Fiocruz), on a suspension of defibrinated and complement-inactivated human blood and parasites in stationary phase of culture growth (Ward et al. 1978), according to the protocols in Table II. The females were examined between 1 and 22 days after the infecting meal. Groups of ten individuals were subjected to 15°C for 3 min and then transferred to micro centrifuge tubes containing sterile PBS (pH 7.2). The intestines were obtained after dissection and examined under a phase contrast microscope (Axioskop, Zeiss) coupled to a photographic system (MC 80, Zeiss). The assessment parameters utilized were: (1) distribution of the flagellates along the digestive tract; (2) degree of infection (+: 1-10 flagellates, ++: 11-30 flagellates, +++: 30-50 flagellates, ++++: uncountable); (3) presence of free or attached forms; (4) blood digestion phases phase 1 (F1): blood retained on the peritrophic matrix, phase 2 (F2): blood partially digested, phase 3 (F3): little blood residue, and phase 4 (F4): absence of blood; (5) infection rate: ratio between the quantity of positive females in F4 (susceptibles) and the total number of dissected females. Parasite isolation and analysis by isoenzyme elec-trophoresis - A total of 77 dissected and examined guts, from females from protocols A.2 (one batch infected by M5725), B.3 and B.4, were inoculated each one (positives or not) in SIM, supplemented with 10% FBS, 200 i.u./ml of penicillin and 50 µg/ml of 5'-fluorocytosine (Sigma). After two-three subcultures, the masses obtained were destined for cryopreservation in SIM, with the addition of 30% FBS and 8% glycerin (Jaffe et al. 1984), and the remainder was prepared for enzyme electrophoresis in agarose gel, in accordance with Franco et al. (1996). As the strains had previously been characterized by means of enzyme activity analysis, isoenzyme electrophoresis and monoclonal antibodies (Medina-Acosta et al. 1994, Franco et al. 1996, 1997a), it was considered that G6PDH (glucose-6-phosphate dehydrogenase, EC 1.1.1.49) system would be sufficient for discriminate between the isoenzyme profile of the samples (Franco et al. 1996). Statistical analysis - Data were analyzed using SPSS for Windows package (10.0.1). Proportions were compared by the chi-square test. Results of independent experiments with primary and cloned samples of E. schaudinni MCHO/BR/79/M5725 were expressed as mean ± standard deviations, and the probability of significant differences between groups was determined by Student's t test. All test were two-tailed and differences at P < 0.05 were regarded as significant. Distribution and infectivity of Endotrypanum spp. in L. longipalpis - Distinct patterns of infections by the strains of Endotrypanum were disclosed. The flagellates of Endotrypanum sp. RO1634 caused abundant infection in the pylorus by attached forms in all tested females (Figure). In the abdominal midgut, flagellates were only seen on the twelfth day post-infecting feeding (p.i.f.), in one female. The infection rate (16/32 females, 50%, Table IV) was significantly greater (P < 0.01) than that caused by E. schaudinni M5725 (16.7%, 4/24 females). Flagellates of E. schaudinni M5725 and c5725 (1 ´ 107) were more abundant (4+) and frequent in the abdominal midgut, pylorus, and ileum from the fifth day p.i.f. onwards, when 80% of the females had already digested the blood. Attached forms could be seen in the pylorus, ileum, and rectal ampulla, with greater intensity and frequency at the first two sites. In the midgut, free flagellates were nearly always observed. At no time there was evidence of infections within Malpighian tubules. Some differences between M5725 and c5725 were recorded: only the primary sample infected the stomodeal valve (1+ in one female, Figure); the c5725 sample infected more females than M5725 (Table III, P < 0.001). In the batches submitted to the lower load (105) of parasites, in one female infected just by c5725 (1/31 females, data not shown), parasites were found in the hindgut (1+), but no female was infected by M5725 (0/28 females). Distribution and infectivity of L. (V.) guyanensis in L. longipalpis - L. guyanensis M4147 was not found in 56 dissected females (Table IV). However, L. guyanensis IM4216, in 11 susceptible flies, showed typically peri-pylarian behaviour: initial infection in the abdominal midgut, while the blood was retained on the peritrophic matrix, followed by escape of the flagellates to the "posterior station" with attachment to the pylorus in large numbers and to a lesser degree in the ileum and rectal ampulla. Migration to anterior station of the intestinal tract was clearly seen in ~55% of females (6/11, Figure) with plentiful infection at the level of the stomodeal valve. Detailed examination to search infection in the Malpighian tubules and in salivary glands could not demonstrate the presence of parasites. Co-infection of L. longipalpis and biochemical analysis of isolates - The results showed a negative re-lation of concomitant infections including L. guyanensis M4147 and the infection rate. That is, despite this parasite had not established in L. longipalpis, the infection rate in co-infected females was reduced drastically, in comparison with single infections with E. schaudinni. We could find infection only in the batch exposed to the mixture with c5725 (Table IV). The distribution of flagellates in the females exposed to the concomitant infection by L. guyanensis IM4216/E. schaudinni M5725 was similar to pure infection by L. guyanensis (data not shown). In particular, susceptible flies displayed migration of flagellates to the foregut: 5 out 12 flies (41.7%) from protocol B.3, and in 5 out of 11 females (45.5%) from protocol B.4, harboured a massive colonization near the stomodeal valve. In one female (protocol B.3), 22 days after the infecting meal, flagellates were found to be plentifully infecting that region. There was no statistically significant difference between the co-infected and L. guyanensis IM4216 single-infected females (Table IV), independently of the proportion of parasites in the mixture. A total of 21 isolates out of 77 guts (27.3% of efficiency) were obtained, from which three (3.9%) were from negative females: (a) 12 positive cultures were from females exposed to E. schaudinni M5725 (protocol A.2), including two from negative specimens by microscopy; (b) six positive cultures including one from negative female (by microscopy) were from 25 guts of females exposed to 1:1 mixture of E. schaudinni M5725/L. guyanensis IM4216 (protocol B.3); (c) three isolates were obtained from guts of four females subjected to 1:19 mixture (protocol B.4); (d) eleven positive cultures from 23 females exposed to L. guyanensis IM4216 were, initially, yielded (one from negative gut by microscopy), but the expansion of cultures failed (not by contamination). The electrophoretic mobilities of enzymes, from all isolates examined, corresponded to E. schaudinni. The results obtained by us and by Franco et al. (1997b) indicate that strains of E. schaudinni present different patterns of behavior in L. longipalpis. Could these differences be related to the large biochemical and molecular heterogeneity found in this genus (Franco et al. 2000)? Our results have also shown that the clone of E. schaudinni infected more females than the primary sample. This can make us formulate the hypotheses that, in the primary sample, are there "subpopulations"? Handman et al. (1983) showed the possibility of the existence of intra-populational heterogeneity in Leishmania tropica, in a study using infective and non-infective clones to ver-tebrate cells. The infections made with Endotrypanum of culture do not produce invasion in Malpighian tubules of L. longipalpis. This conclusion confirms the results of Brazil et al. (1991) e Franco et al. (1997b). Shaw (1981), on the other hand, fed females of L. longipalpis directly in a sloth naturally infected with E. schaudinni M5725 and evidenced infections in the tubules. Possibly it has happened, after long subcultivation, lineage selection of parasites that adapted themselves to the axenic medium where pH is alkaline. The subpopulation with capacity to invade the Malpighian tubules, which has an acid pH (Gontijo et al. 1998), could have been eliminated or reduced. The scarcity of Endotrypanum infections in the region near the stomodeal valve leads us to a polemic issue. First, it is important to consider that this evidence was also obtained in other works (Shaw 1981, Brazil et al. 1991). It is very significant that Shaw (1981) had started the female infections of L. longipalpis and L. flaviscutellta with the forms found in the vertebrate host and had not found flagellates in the above-mentioned region. On the other side, Shaw (1964), in L. sanguinaria and L. trapidoi, and Christensen and Herrer (1976), in these same species and in L. gomezi, and Shaw (1981), in L. furcata (1 female only), reported infections in the stomodeal valve level. Were the flagellates seen by these authors really Endotrypanum? Zeledón et al. (1979) argued that it would be possible that some of the infections reported by Shaw (1964) and Christensen and Herrer (1976) might have been due to L. herreri sp. The distribution of L. (V.) guyanensis MHOM/BR/95/IM4216 followed the pattern typical of Leishmania (V.) spp. (Rangel et al. 1992, 1993). In the present work, not only were flagellates found in a large percentage of susceptible females, but they were also seen to be abundantly infecting the stomodeal valve. Thus, the data achieved here suggest that the distribution of Endo-trypanum spp. in L. longipalpis from the Lapinha Cave showed limited similarity to the distribution of L. (V.) guyanensis. In spite of the records of Endotrypanum and Leishmania sharing the digestive tract of sand flies, so far there have not been studies about selection using these trypanosomatids. We have shown evidences which suggest that the presence of L. guyanensis M4147 (non-infective) presumably inhibited the development of Endotrypanum, with the drastic reduction in the number of positive females, mainly in the group of females exposed to coinfection with the heterogeneous sample (M5725) of E. schaudinni. Effects of intra- or interspecific selective pressures have already been reported previously, among co-cultivated trypano-somatids (Pacheco et al. 1987, Coppens et al. 1992) and even about the biology of sand flies (Alekseev et al. 1975, El-Sawaf et al. 1994). On the other hand, the correlation between the massive infection found in the stomodeal valve of females coinfected with L. guyanensis IM4216 and the isoenzymatic char-acterization of isolates suggest that there had been selection in the culture medium. Deane et al. (1984) demonstrate that the methods of parasite isolation and further passages in culture may completely eliminate one strain from originally mixed population. So, the electrophoretic profiles of isoenzymes of parasites recovered from females submitted to co-infections, including flagellates from negative females by optical microscopy suggest that the mere correlation of sand fly infection/zymodeme may induce parasitological and epidemiological incorrect considerations and that the natural infections, single or concomitant, of sand flies by Endotrypanum and Leishmania are underestimated. These questions lead us to the importance of doing further studies on the interaction between these trypa-nosomatids and sand flies, specially using a marker for selection inside the sandfly gut and molecular methods to identify the isolates with sensitivity and specificity. To Cláudia A Gonçalves (Departamento de Entomologia, IOC-Fiocruz), for expertise in rearing L. longipalpis. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06140t3.jpg] [oc06140t1.jpg] [oc06140f1.jpg] [oc06140t4.jpg] [oc06140t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}