 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101, No. 8, December ,2006, pp. 917-921 Higher genetic variation estimated by microsatellites compared to isoenzyme markers in Aedes aegypti from Rio de Janeiro Magda Clara Vieira da Costa-Ribeiro, Ricardo Lourenço-de-Oliveira/+, Anna-Bella Failloux*/++ Laboratório
de Transmissores de Hematozoários, Instituto Oswaldo Cruz-Fiocruz,
Av. Brasil 4365, 21040-900 Rio de Janeiro, RJ, Brasil *Insectes
et Maladies Infectieuses, Institut Pasteur, Paris, France Finnancial suport: Pasteur Institute, CNPq (Proc. 306111/ 2003-9), Capes Received
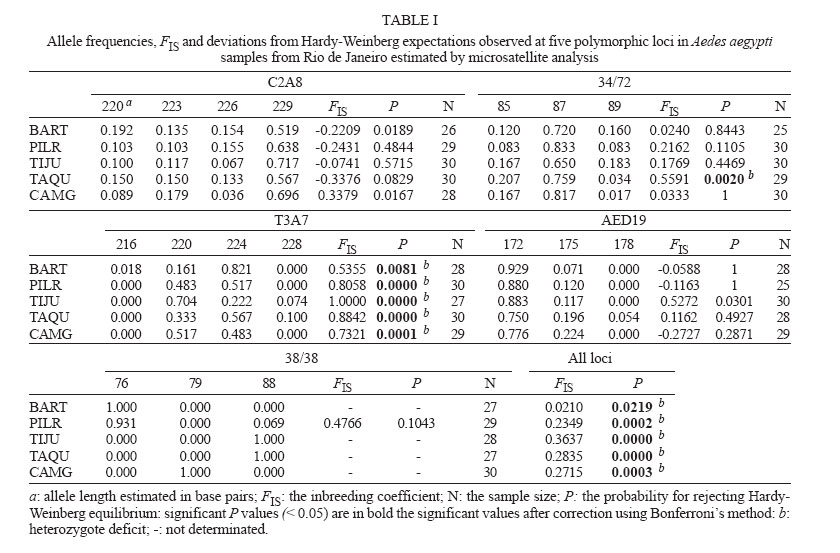
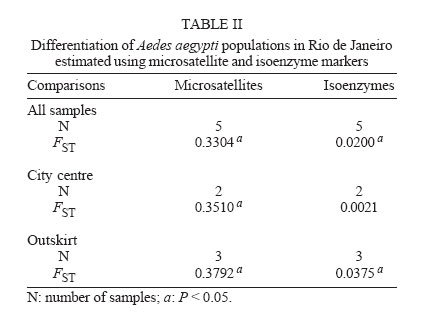
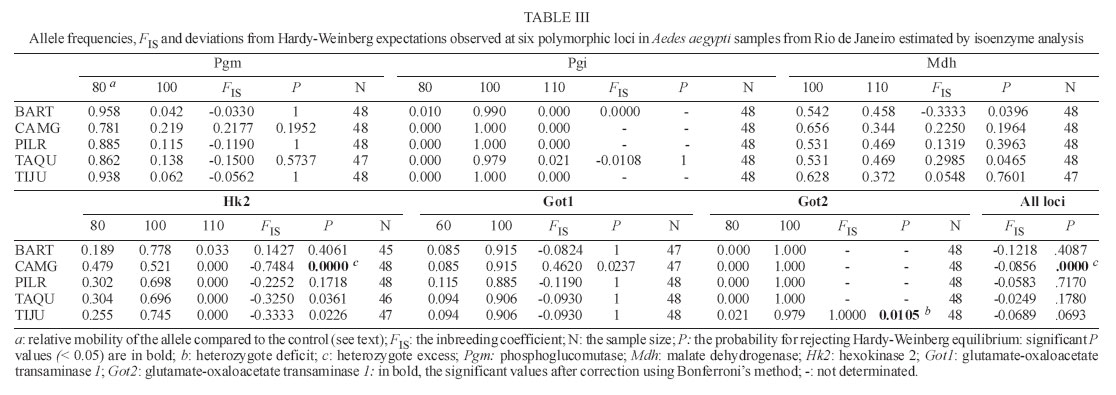
11 July 2006 Code Number: oc06150 Aedes aegypti populations from five districts in Rio de Janeiro were analyzed using five microsatellites and six isoenzyme markers, to assess the amount of variation and patterns of gene flow at local levels. Microsatellite loci were polymorphic enough to detect genetic differentiation of populations collected at small geographic scales (e.g. within a city). Ae. aegypti populations were highly differentiated as well in the city center as in the outskirt. Thus, dengue virus propagation by mosquitoes could be as efficient in the urban area as in the outskirt of Rio de Janeiro, the main entry point of dengue in Brazil. Key words: Aedes aegypti - microsatellites - isoenzymes - genetic differentiation Dengue is the most important mosquito-borne disease in Brazil where it is essentially transmitted by Aedes aegypti. Rio de Janeiro is considered to be the main introduction location for dengue viruses which have caused most epidemics in Brazil with increasing incidence of severe cases (Nogueira et al. 2002, Teixeira et al. 2002). The urban mosquito Ae. aegypti is a major dengue vector in Brazil. It is present in high density at the vicinity of highly populated areas (Lourenço-de-Oliveira et al. 2004). The species shows high level of resistance to insecticides (Luz et al. 2003, Braga et al. 2004, Da-Cunha et al. 2005) and is highly susceptible to dengue viruses (Lourenço-de-Oliveira et al. 2004). Moreover, uncontrolled urbanization and intensive travel and migration are additional factors that favor both vector and virus dis-semination (Gubler 1998, Herrera et al. 2006). Studies on Ae. aegypti population structure based on allozyme analysis have been reported for more than 40 years (Tabachnick et al. 1979). More recently, micro-satellite markers have been extensively applied to study genetic variations or to detect gene flow (Bruford & Wayne 1993, Jarne & Lagoda 1996). Microsatellites are apparently neutral, codominant and highly polymorphic markers (Tautz 1989). Field studies on genetic differentiation within vector populations can yield important information regarding to evolution and population biology (Wang et al. 2001). Variations in isoenzymes and microsatellites among vector populations have provided evidence that water storage habits and human densities may affect their differentiation (Huber et al. 2002). Besides, micro-satellite markers provide a sensitive measure of divergence and therefore can potentially distinguish populations that may have recently diverged (Lanzaro et al. 1995). Here, we report a genetic study on Ae. aegypti in Rio de Janeiro using both microsatellite and iso-enzyme markers. MATERIALS AND METHODS Mosquito samples - Ae. aegypti was sampled in five districts in Rio de Janeiro in March 2003. Samples were divided into two groups according to the human density in the sampled district: (i) high human density (city center) Pilares (PILR) and Tijuca (TIJU) (15,364.17 inhabitants/km² and 17,431.88 ha/km2, respectively) and (ii) moderate human density (outskirt) - Barra da Tijuca (BART), Campo Grande (CAMG), and Taquara (TAQU) (1,726.46 ha/km2, 2,182.58 ha/km2 and 6,444.13 ha/km2, respectively). To avoid collections of descendents from small number of Ae. aegypti females, 20 ovitraps (Reiter et al. 1991) were set per district during two consecutive weeks (Dibo et al. 2005). Districts are separated by a minimal distance of 8.2 km and a maximal of 32.4 km (see Costa-Ribeiro et al. 2006 for more details). Mosquitoes were reared to the adult stage (F0 gen-eration) in insectaries under standardized conditions (25 ± 1°C, 80 ± 10% relative humidity and 12 h light/dark cycle) and subsequently, stored at 80°C. Mosquitoes from the same samples were analyzed by both the microsatellite and isoenzyme assays. Microsatellites - Thirty mosquito adults from each district were analyzed using six microsatellite loci: C2A8, 34/72, T3A7, AED19, 38/38, and 9A89 (see Huber et al. 2001 for more details). DNA was extracted in DNAzol solution (Gibco BRL). PCR reactions containing 1X buffer (Eurobio), 1.2 mM MgCl2, 60 µM of each dNTP, 5 pmol of each primer, 0.25 units of Taq polymerase (Eurobio), and 2 µl of DNA were performed in a 9600 thermal cycler (Perkin-Elmer). The reaction conditions were 5 cycles (2 min at 96°C, 30 s annealing at locus-specific, an-nealing temperature (Ta) and 1 min 15 s extension at 72°C) followed by 25 cycles (30 s at 95°C, 30 s at Ta and 1 min 15 s at 72°C) and a final elongation step (5 min at 72°C). One primer (1.5 pmoles) was end-labeled with one of the three different fluorescent phospho-ramidite dyes (FAM, HEX or NED) appropriate for ABI PRISM instruments. One microliter of each PCR reaction was diluted to 1:20 times with distilled water. Samples were prepared by adding 0.5 µl of internal size standard 400HD ROX (Perkin Elmer) and deionised formamide for a final volume of 20 µl. Isoenzymes - Forty-eight adults from each district were analyzed for 10 enzyme systems: glucose phosphate isomerase (Gpi, EC 5.3.1.9.), glutamate oxaloacetate transaminases (Got1 and Got2, EC 2.6.1.1.), glycerol phosphate dehydrogenase (Gpd, EC 1.1.1.8.), hexokinases (Hk, EC 2.7.1.1.), malate dehydrogenase (Mdh, EC 1.1.1.37.), malic enzyme (Me, EC 1.1.1.40.), and phosphoglucomutase (Pgm, EC 2.7.5.1.). Each individual mosquito was ground in 25 µl of distilled water and centrifuged (12,000 g for 3 min at +4°C). The supernatant containing soluble proteins was loaded onto a 12.8% starch gel in Tris-Maleate-EDTA (pH 7.4) buffer and run for 4-5 h (Pasteur et al. 1988). A laboratory strain of Ae. aegypti named "Paea" (collected in 1994 in Tahiti, French Polynesia) was used as a mobility control for isoenzyme polymorphism. Genetic analysis - Hardy-Weinberg (HW) propor-tions were compared using GENEPOP software (version 3.4) (Raymond & Rousset 1995). Deviations from HW were tested using an exact test procedure with the alternative hypothesis H1 corresponding to deficits or excess in heterozygotes (Rousset & Raymond 1995). Linkage disequilibrium was tested between pairs of loci for each sample using Fisher's exact test on rank ´ column contingency tables. FIS, the inbreeding coef-ficient, and FST, the fixation index, were estimated as described by Weir and Cockerham (1984). Genetic differentiation across samples was estimated by calculating the P value associated to the FST estimate. Significance levels for multiple testing were corrected using sequential Bonferroni's procedures (Holm 1979). Genetic isolation by geographic distance was tested by estimating rank correlations between FST/(1-FST) calculated between pairs of samples and Ln distances (Slatkin 1993). RESULTS Microsatellite polymorphism - The locus 9A89 was monomorphic and 3-4 alleles were scored for the other five loci: three for 34/72, AED19, and 38/38, and four for C2A8 and T3A7 (Table I). The allelic frequencies varied for the loci C2A8 from 0.036 (allele 226) in CAMG to 0.717 (allele 229) in TIJU; for 34/72, from 0.017 (allele 89) in CAMG to 0.833 (allele 87) in PILR; for T3A7, from 0.018 (alleles 216) to 0.821 (allele 224) in BART; for AED19, from 0.054 (allele 178) in TAQU to 0.929 (allele 172) in BART; and for 38/38, from 0 to 1 (Table I). For HW equilibrium, 90 tests were carried out (1 test for each locus in each sample). Significant deviations from Hardy-Weinberg equilibrium associated to deficits of heterozygotes were detected in the locus 34/72 in TAQU (FIS = 0.5591, P = 0.0020) and in the locus T3A7 for all samples: BART (FIS = 0.5355, P = 0.0081), PILR (FIS = 0.8058, P <10-4), TIJU (FIS = 1, P < 10-4), TAQU (FIS = 0.8842, P < 10-4), and CAMG (FIS = 0.7321, P = 0.0001). Tests considering all loci for each sample showed significant deviations from Hardy-Weinberg equilibrium due to heterozygote deficits (Table I). No linkage disequilibrium was detected among 34 comparisons tested. Genetic differentiation evaluated by estimating FST values was high and significant when considering mosquitoes from all five districts (FST = 0.3304, P < 0.05; Table II). When samples were grouped according to human density (city center versus outskirt), a higher genetic differentiation (FST = 0.3792, P < 0.05) was detected among samples from the outskirt (Table II). When testing isolation by geographic distance by estimating rank correlations between FST/(1-FST) calculated between pairs of samples and log geographical distances, no isolation was demonstrated (P = 0.1522) (data not shown). Isoenzyme polymorphism - Six (Pgm, Pgi, Mdh, Hk2, Got1, and Got2) out of 10 loci investigated were poly-morphic. 2-3 alleles were scored for each locus: 2 for Pgm, Mdh, Got1 and Got2, and 3 for Pgi and Hk2 (Table III). The allelic frequencies varied for the loci Pgm from 0.115 (100) in PILR to 0.958 (80) in BART; for Mdh, from 0.344 (110) in CAMG to 0.656 (100) in CAMG; for Hk2, from 0 to 0.778 (100) in BART; for Got1, from 0.085 (60) in CAMG to 0.915 (100) in BART and CAMG; and for Pgi and Got2, from 0 to 1 (Table III). For Hardy-Weinberg equilibrium, 150 tests were carried out. Only two significant deviations from Hardy-Weinberg equilibrium were observed: a heterozygote deficit in TIJU (FIS = 1, P = 0.0105) and a heterozygote excess in CAMG (FIS = -0.7484, P < 10-4 (Table III). When considering all loci in each sample, a significant heterozygote excess was observed in CAMG (FIS = -0.0856 and P < 10-4) (Table III). No linkage dis-equilibrium was detected among 42 comparisons. Differentiation was significant when considering all samples (FST = 0.0200, P < 0.05), and samples from the outskirt (FST = 0.0375, P < 0.05) (Table III). No correlation was detected when testing genetic isolation by geographic distance (data not shown). As expected, microsatellite markers displayed higher polymorphism when compared to isoenzyme loci. For microsatellite loci, five among six loci were polymorphic whereas for isoenzyme loci, only six among ten were polymorphic. Microsatellite loci in Ae. aegypti populations from Rio de Janeiro were less polymorphic when compared with Asian populations (Huber et al. 2002, 2004, Paupy et al. 2004). Low abundance and limited polymorphism in microsatellites have been already described for Ae. aegypti (Fagerberg et al. 2001, Ravel et al. 2001, 2002). In all samples, heterozygote deficits were observed due to the locus T3A7. This may be related to four main factors: (i) the locus is under selection, (ii) null alleles may be present, (iii) inbreeding may be common in populations, or (iv) the presence of population substructure which leads to a Wahlund's effect. The locus T3A7 was probably subjected to mutations in its flanking regions which resulted in inhibition of primer annealing with the complementary DNA sequences. The failure in detection of microsatellite PCR products generates null alleles and thus, a deficit in heterozygotes (Lanzaro et al. 1995). Inbreeding could not explain heterozygote deficits as deficits were not observed in all loci. The Wahlund effect which describes deviations due to the pooling of subpopulations in HW equilibrium could be one explanation. However, the less polymorphic isoenzyme loci only detected two samples with a deviation from HW equilibrium due to a heterozygote excess or deficit. Isoenzyme markers revealed a larger genetic dif-ferentiation among samples from the outskirt whereas samples from the city center were not differentiated. This pattern contrasted with those observed in Vietnam (Tran Khanh et al. 1999). Ae. aegypti females tend to lay eggs in artificial containers at the vicinity of human habitations. So, dispersal varies upon the distribution of breeding sites and human density for blood feeding (Tsuda et al. 2001). Gene flow contributes to homogenize populations and thus reduces genetic differences among them. In Rio de Janeiro, samples were collected at the end of the rainy season, in March, when a large number of unintentional containers such as cans, tires and bottles were productive in larvae and pupae (Honório et al 2006). This type of breeding sites more abundant in highly populated areas predominates in the city center where flow low genetic differentiation or deficits among mosquito population was detected using isoenzyme markers. Conversely, microsatellite markers were polymorphic enough to differentiate populations within the city center whereas isoenzymes failed to detect any differentiation. Dengue is endemic in Rio de Janeiro since the in-troduction of dengue virus in 1986 (Lourenço-de-Oliveira et al. 2004). Variations in dengue incidence are observed within the city depending on vector distribution and vector control implementation (Luz et al. 2003). Our results demonstrated microsatellite markers to be more adapted to evaluate genetic structure of samples collected within a small geographic scale like a city. The same pattern of genetic differentiation of Ae. aegypti populations in the city center and the outskirt suggested that the dengue virus propagation by mosquitoes could be as efficient in urban area as in the outskirt of Rio de Janeiro. To Mauro Blanco Brandolini, Fábio Castelo, Fernando da Costa Alves, and the staff of Coordenação de Controle de Vetores in Rio de Janeiro for providing mosquito samples; to Nadia Ayad for technical assistance; to Sara Moutailler and Marcia Castro for their assistance in the laboratory. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06150t3.jpg] [oc06150t1.jpg] [oc06150t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}