 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101,Suppl. I, October ,2006, pp. 137-143 Protein tyrosine kinases in Schistosoma mansoni Diana Bahia/+, Luiza Freire Andrade*, Fernanda Ludolf*, Renato Arruda Mortara**, Guilherme Oliveira/* Centro de Pesquisas René Rachou-Fiocruz, Av. Augusto
de Lima 1715, 30190-002 Belo Horizonte, MG, Brasil *Programa de Pós-Graduação
e Pesquisa, Santa Casa de Misericórdia de Belo Horizonte, Belo Horizonte,
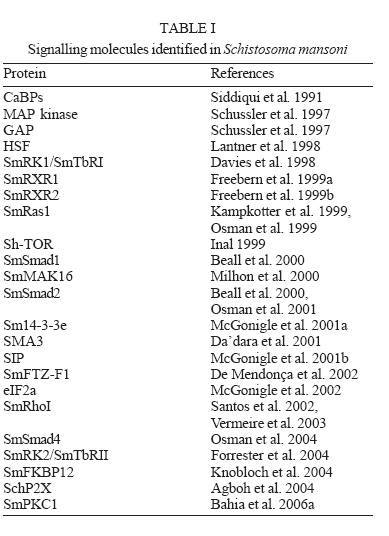
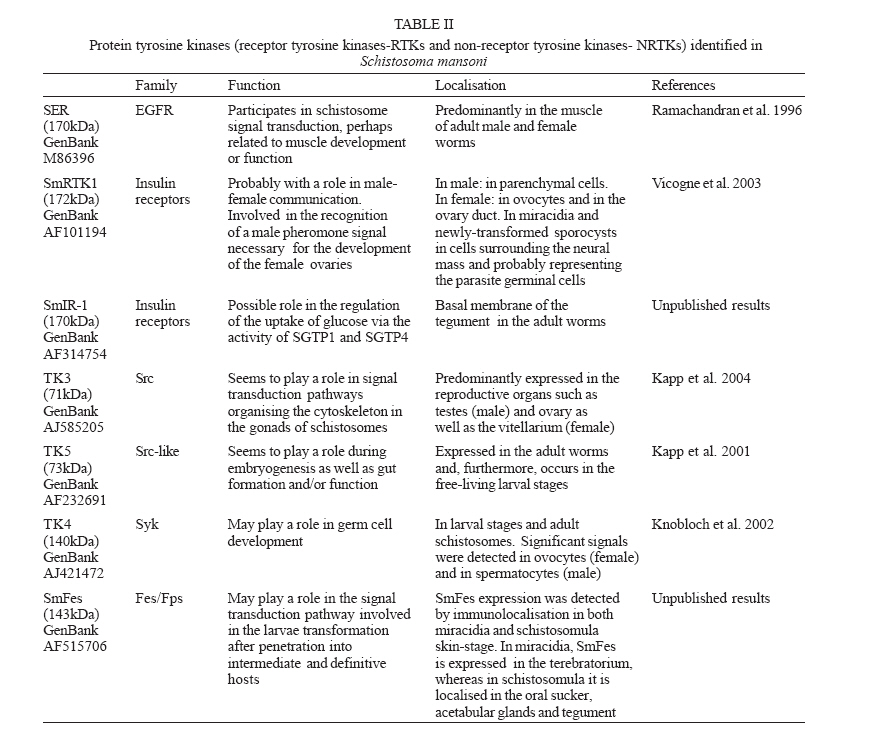
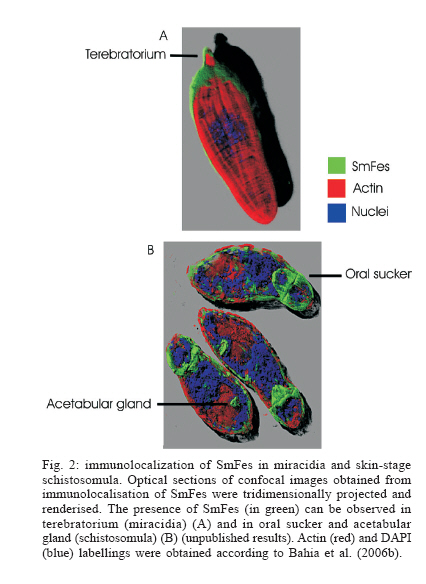
MG, Brasil Financial support: Fapemig (DB CBB-174/02) Received 25 May 2006 Code Number: oc06180 The identification and description of signal transduction molecules and mechanisms are essential to elucidate Schistosoma mansoni host-parasite interactions and parasite biology. This mini review focuses on recent advancements in the study of signalling molecules and transduction mechanisms in S. mansoni, drawing special attention to the recently identified and characterised protein tyrosine kinases of S. mansoni. Key words: signal transduction - protein kinase - schistosoma - tyrosine kinase The identification and characterisation of signal transduction molecules and mechanisms are essential to elucidate Schistosoma mansoni host-parasite interactions and parasite biology. Protein tyrosine kinases (PTKs) are important molecules for intra- and inter-cellular communication as well as for survival in eukaryotes, playing a major role in signal transduction processes (Hanks et al. 1988). PTKs also participate in cellular mechanisms that control several biological processes such as adhesion, cytoskeleton reorganisation, and migration. These proteins are also known to be involved in developmental and differentiation processes of cells. Therefore, the study of PTKs may unveil strategies that can be used for identifying new candidate drug targets. Phosphorylation of protein substrates catalysed by kinases is an essential mechanism by which important intracellular and extracellular signals are transmitted throughout the cell and to the nucleus (Cheetham 2004). During kinase catalysis, the g-phosphate group from ATP is transferred to the protein substrate, thereby changing the substrate properties (e.g. its structure, location or its activity as enzyme) (Manning et al. 2002a). The recent success in cancer treatment that includes specific tyrosine kinase inhibitors strongly validates the clinical relevance of basic research on tyrosine phosphorylation. Functional profiling of the tyrosine phospho-proteome is likely to lead to the identification of novel targets for drug discovery, providing a basis for novel molecular target approaches (Machida et al. 2003). Many of the 500 or so identified human protein kinases are attractive drug targets to treat cancer, inflammation processes and autoimmune diseases. PTKs PTKs comprise proteins found in multicellular organisms (Neet & Hunter 1996). PTKs may be found (i) anchored in the cell membrane, acting as receptors, (ii) free in the cytoplasm, participating in signalling cascades, and (iii) in the nucleus, directly associated with gene activation processes. PTK catalytic domain activation results from the interaction with other signalling proteins, which allows specifically signalling propagation. There is no evidence of PTK in yeast. However, 49 from 239 Drosophila melanogaster PKs and 105 out of 454 Caeno-rhabditis elegans PKs have been classified as PTKs (Manning et al. 2002a). There are two major classes of PTKs, receptor tyrosine kinases (RTKs) and non-receptor tyrosine kinases, also named cytoplasmic or cellular tyrosine kinases (NRTKs) (Neet & Hunter 1996). Receptor tyrosine kinases (RTKs) - RTKs contain three distinct regions: an extracellular binding domain, a transmembrane helix, and a cytoplasmatic domain that contains the kinase activity (Hubbard & Till 2000). RTK activation is generally triggered by the interaction of a ligand with a specific biding site on the receptor extracellular domain. Following RTK stimulation at the extracellular site, the catalytic domain on the cytoplasmic side of membrane is activated by the dimerisation of the receptor, leading to autophosphorylation (Alberts et al. 1994) and then to the activation of kinase activity, providing a new biding site for intracellular adapter molecules. RTK families show diversified extracellular domains (Heldin 1996). The first protein receptor described as being protein kinase tyrosine-specific was the epidermal growth factor receptor (EGFr) (Yarden & Ullrich 1988). However, there are growth and differentiation factor receptors that also belong to RTKs. Non-RTKs (NRTK) - NRTKs are a set of intracellular signalling proteins that has been identified as being able to interact with PTK phosphotyrosine (Neet & Hunter 1996). Although intracellular signalling proteins that bind to activated PTK phosphotyrosine residues have various functions and structures, they generally share highly conserved non-catalytic domains, known as SH2 and SH3. One of the main features of NRTK is the presence of the SH2 domain. SH2 domains are small protein modules that bind specifically to tyrosine-phosphorylated peptides. There are more than 100 SH2 domains in the human genome, and different SH2 domains bind to different classes of tyrosine-phosphorylated ligands. These domains play a critical role in the propagation of signals in the cell, mediating the relocation and complex formation of proteins in response to changes in tyrosine phosphorylation (Machida et al. 2003). Protein kinases and schistosome biology Animal cells normally divide when stimulated by growth factors, which are generally produced by other cells and act via RTKs. Given that mutations leading to amino acid substitutions on protein kinases are common in cancer, diabetes and other diseases, a better understanding on how such enzymes regulate a wide range of functions may enable further therapeutic interventions (Plowman et al. 1999). Further knowledge on PTKs may provide new strategies for drug development, an approach intensively pursued in cancer research (Traxler 2003, Harari 2004). As sensing and responding to the environment are essential in the complex life cycle of schistosomes, the knowledge gained by studying signal transduction proteins and their mechanisms will be important for understanding the biology of the organism. S. mansoni proteins involved in signal transduction It is now clear that the ability of S. mansoni to survive for decades in the blood-stream of its host and the sexual maturation of the female that depends on a close contact with the male are processes that require molecular communication (Schussler et al. 1997, Kunz 2001, Kapp et al. 2004). Recently, a number of signalling molecules have been identified and cloned in schistosomes, including transmembrane and cellular receptors (Table I). Some of the signalling proteins seem to be involved with the SmRK (S. mansoni receptor kinase) signalling pathway. SmRK is a divergent member of the serin/threonin kinase TGF-b receptor (transforming growth factor beta receptor family), possibly participating in the host response to growth factors such as: cell migration, differentiation, adhesion and apoptosis. SmRK1 is a surface membrane receptor serine/threonine (Davies et al. 1998) belonging to the S. mansoni TGF-b superfamily that may be important in mediating host-parasite interactions associated with parasite development. Sm14-3-3e is a cytoplasmatic protein associated with TGF-b (McGonigle et al. 2001a). The overexpression of Sm14-3-3a leads to increased TGF-b signalling, whereas eIF2a (eukaryotic initiation factor 2 alpha subunit) leads to TGF-b inhibition (McGonigle et al. 2002). Smads are able to interact with receptor molecules carrying the message to the nucleus. Both Smads (SmSmad1 and SmSmad2) and SmRK1 are found in the same developmental stages (lung stage and adult parasites). SmSmad2 interacts with SmRK1, while SmSmad4 interacts with SmSmad1 and SmSmad2, besides phosphorylating Erk1/2 (kinase regulated by extracellular signal) (Beall et al. 2000, Beall & Pearce 2001, Osman et al. 2001, 2004). FKBP12 influences a variety of signal transduction pathways that regulate cell division, differentiation, and ion homeostasis. Among these, TGF-b signalling and calcineurin (CN) phosphatase activity are modulated by FKBP12 via binding to TGF-b family type I receptors (TGFbR-I) or to the CN subunit A, respectively (Chen et al. 1997). The S. mansoni FKBP12 homologue (Sm-FKBP12) is a direct partner of SmRK1 and both are present and interact in the female gonads (Knobloch et al. 2004). Other signalling proteins found in S. mansoni participate in several different pathways and functions. SmPKC1 has been recently described by our group as the first b1- type protein kinase C identified in S. mansoni. Immuno-localisation studies indicated that SmPKC1 was strongly associated with the ridge cyton and excretory vesicles in sporocysts, while in skin-stage schistosomula, SmPKC1 was clearly expressed in the acetabular gland, tegument, and duct (Bahia et al. 2006a). SmMAK16 contains a nuclear signalling portion and a site for CK2 (casein kinase 2) phosphorylation, being related to the biogenesis of the ribosome 60S subunit as well as to the cell cycle with higher expression levels in female worms (Milhon et al. 2000). SmRXR are nuclear receptors and gene transcription activators. The SmRXR gene is constitutively expressed and thus must play multiple roles throughout the schistosome life cycle. SmRXR is located in vitellinic cells, and may play a role in the activation of the eggshell p14 gene precursor. (Freebern et al. 1999a,b, Fantappié et al. 2001). SmFTZF1 is another nuclear receptor with a highly conserved DNA binding domain, related with developmental and sexual differentiation (de Mendonça et al. 2002). SchP2X is related to the ATP ionic channel opening (Agboh et al. 2004). SmRhoI (Santos et al. 2002, Vermeire et al. 2003) is a GTPase that possibly participates in the cytoskeleton organisation, gene transcription, cell cycle and membrane transport and is expressed at higher levels in female worms (Vermeire et al. 2003). SMA3 is a Ca-ATPase homologue found in the adult tegument, suggesting that SMA3 functions to help control Ca homeostasis within the tegument and may play a role in signal transduction at the host-parasite interface (Da'dara et al. 2001). PTKs identified in S. mansoni Very few PTKs have been identified and characterised in S. mansoni: three of them are RTKs (SmRTK-1, SmIR-1, SER), and four are NRTKs (TK5, TK4, TK3, SmFes), (Table II). SmRTK1 is a membrane protein with an extracellular binding domain similar to several protein domains that share the Venus Flytap-VFT structure and the cytoplas-matic TK domain, which is similar to the insulin receptor (IR) catalytic domain. The SmRTK1 gene is expressed throughout all developmental stages. In males, it is preferably found in parenchyma cells. In females, an intense labelling was associated with ovocytes present in the ovary and in the ovary duct. SmRTK1 is believed to constitute an original GABA-activated RTK, which is involved in pheromone recognition, necessary for the development of the female ovaries (Vicogne et al. 2003). SmRTK1 was localised in sporocysts. The preferential localisation of SmRTK1 in sporocysts germinal cells and ovocytes could point to a role in schistosome growth and differentiation. SmIR-1 is a tyrosine kinase similar to the family members of IR. It has all the features of IR with a conserved ligand-binding domain. Immunohistochemical studies have shown that SmIR-1 is mainly expressed at the basal membrane level of the tegument in adult worms (Dissous et al. 2006, Khayath et al. unpublished results). It might play a role in glucose uptake regulation. SER is an epithelial growth factor receptor that contains a TK domain homologous to the TK domain of the erbB family. The gene is translated into a 170 kDa protein that contains a signal peptide, a cystein-enriched extracellular domain, a transmembrane hydrophobic sequence, and an intracellular TK domain. The SER protein is present in cercariae and, more strongly, in adult worm muscles (Ramachandran et al. 1996), suggesting that it could participate in muscle development and function. The genes produce three variant transcripts, resulting from SER alternative splicing (Shoemaker et al. 1992). SER seems to be activated by vertebrate EGF ligands besides activating ERK signalling, suggesting a conservation of the EGFR function in Schistosoma (Vicogne et al. 2004). The NRTKs identified in S. mansoni have common domains, as shown in Fig. 1. TK5 and TK3 (Fig. 1, Table II) are NRTK orthologues to the Src family. They contain SH3 and SH2 domains followed by a C-terminal catalytic domain with a tyrosine kinase activity. TK5 is the first Fyn subfamily identified in invertebrates (Kapp et al. 2001). TK3 is a single-copy gene and it codes for a 71 kDa protein expressed in adult worms of both sexes, predominantly in the reproductive organs. Its enzymatic activity was experimentally demonstrated in a heterologous eukaryote cell culture system, which was able to phosphorylate p130Cas, a protein closely related to proteins involved in focal adhesion and cytoeskletal organization (Kapp et al. 2004). Most Src family kinases contain an N-terminal Met-Gly-Cys consensus sequence that undergoes dual acylation with myristate or palmitate after methionine removal (Pellman et al. 1985, Resh 1994). Although this has not been yet experimentally demonstrated, both TK3 and TK5 presumabily have an N-terminal my-ristylation motif, responsible for targeting the Src-related proteins to cellular membranes. TK4 (Fig. 1,Table II) is a NRTK orthologue to the Syk family. TK4 contains two SH2 domains and one tyrosine kinase domain. It is transcribed in maturing and mature adult stages, as well as in larval stages miracidia and cercariae. The presence of TK4 in oocysts and spermatocytes suggests that this protein plays a role in germinative cell development. However, there are no report in literature on the involvement of Syk in gonad differentiation (Knobloch et al. 2002). Our group has previously identified and characterised the molecular structure of a new PTK in S. mansoni, SmFes Fig. 1, Table II, Ludolf et al. unpublished results). SmFes exhibits the characteristic features of Fes/fps protein tyrosine kinases subfamily, a coiled-coil region, SH2 and protein tyrosine kinase catalytic domain signatures, but lacks the FCH amino-terminal domain. It is the first member of Fes subfamily described in helminths. SmFes expression was detected by immunolocalisation in both miracidia and skin-stage schistosomula (Bahia et al. unpublished results). In miracidia, SmFes is expressed in the terebratorium (Fig. 2a), whereas in schistosomula it is localized in the oral sucker, acetabular glands, and tegument (Fig. 2b). In cercariae, an expression pattern similar to schistosomula was observed (not shown). Due to functions attributed to both acetabular glands, oral sucker and terebratorium, these findings seem to suggest that SmFes plays a pivotal role in the signal transduction pathway involved in the larvae transformation after penetration into intermediate and definitive hosts. Presence of PKs in sequenced genomes PKs have been characterised not only through traditional biochemical techniques but also by catalytic domain analyses from aminoacid sequences of their primary structure (Hanks et al. 1988). PKs comprise one of the largest families of proteins, which correspond to 1.5 to 2.5% of eukaryote genes (Manning et al. 2002a). The development of genomic studies has led to the identification of an increasing number of PK in various animal species. The genome of S. mansoni is currently being assembled and annotated and two large scale transcriptome projects described the majority of the genes of this species (El Sayed et al. 2004, Oliveira et al. 2004, Oliveira & Bahia 2004, Verjovski et al. 2004). It is expected that the release of a full analysis of these projects will reveal a large number of proteins related to parasite-environment and parasite-host interactions, among them PTks. By using C. elegans as model for studies on signal transduction, as it was the first fully sequenced multicellular organism, Pks were grouped into the second biggest family of the protein domains in theses worms, comprising 411 fully sequenced PKs (Plowman et al. 1999). Sequencing of the human genome revealed 518 PK coding genes, 1.7% of the entire human gene content. Among the 258 PKs analysed, 83 domain types were identified, most of which are found to be closely related to signalling protein interaction domains, e.g., SH2, which recognises and binds to phosphorylated tyrosine residues (Manning et al. 2002b). By comparing kinomes from S. cerevisiae (yeast), C. elegans (worm), D. melanogaster (insect) and Homo sapiens (mammal), out of 209 subfamilies analysed, 51 proteins were present in the four genomes, 7 were only present in yeast (S. cerevisiae and S. pombe), 15 in C. elegans, 13 in H. sapiens, and none in D. melanogaster (Manning et al. 2002a). PKs represent promising drug targets for a number of human and animal diseases. The recent completion of the genomes of three human-infective trypanosomatid protozoa, Leishmania major, Trypanosoma brucei and T. cruzi, has allowed the kinome for each parasite to be defined as 179, 156, and 171 eukaryotic protein kinases, respectively (Naula et al. 2005), that is about one third of the human complement. Last but not least, several lines of evidence suggest that it is worth pursuing the study of the PK family to identify drug targets for shistosomes. PKs have been shown to be essential for proliferation and/or viability of parasite life-cycle stages that are clinically relevant. As a general rule, PKs having more than 60% sequence identity over the core catalytic domain have a high probability of being inhibited by the same group of low molecular mass compounds. The corollary is that "there is a higher level of confidence that specific inhibitors can be designed to target protein kinases with < 60% sequence identity, which is the case for the vast majority of parasite kinomes when compared to that of humans" (Vieth et al. 2004, Naula et al. 2005). REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06180f2.jpg] [oc06180t2.jpg] [oc06180t1.jpg] [oc06180f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}