 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101,Suppl. I, October ,2006, pp. 307-312 The schistosome enzyme that activates oxamniquine has the characteristics of a sulfotransferase Livia Pica-Mattoccia, Daniele Carlini, Alessandra Guidi, Velasco Cimica, Fabio Vigorosi, Donato Cioli+ Institute of Cell Biology, CNR, 32 via Ramarini, 00016 Monterotondo, Rome,
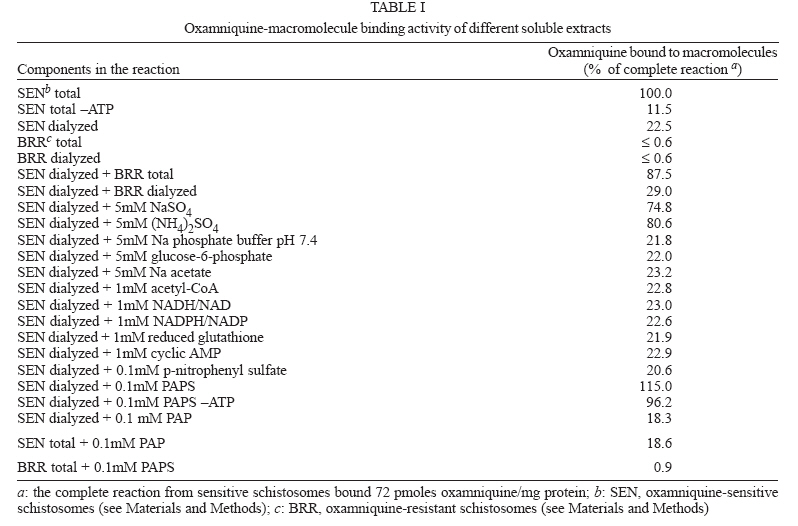
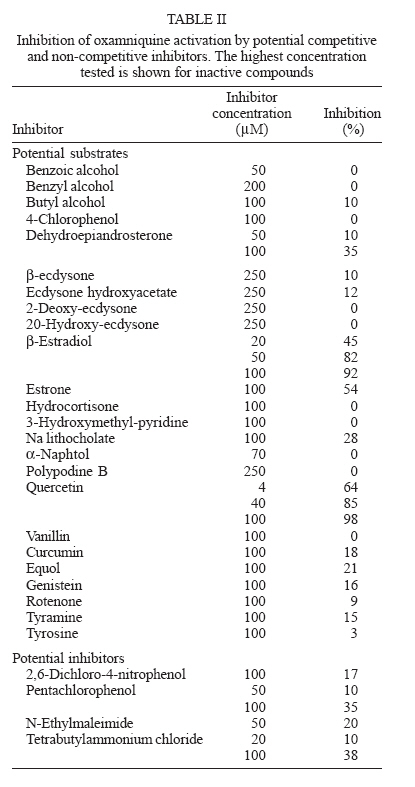
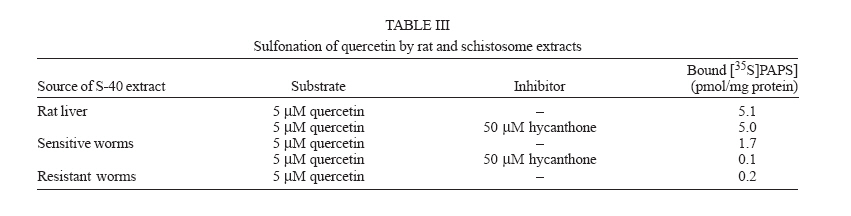
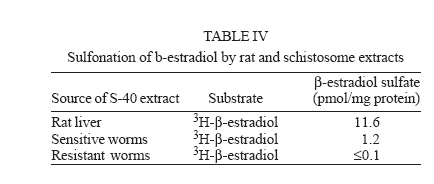
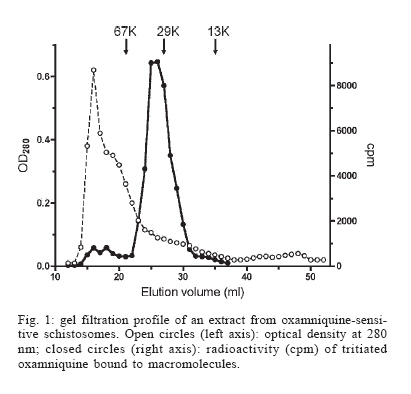
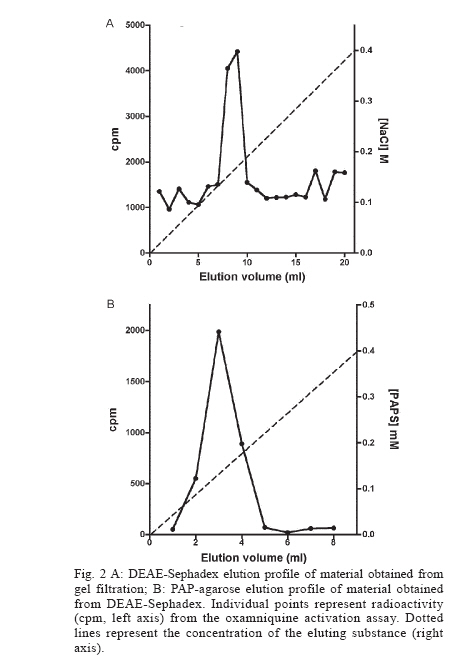
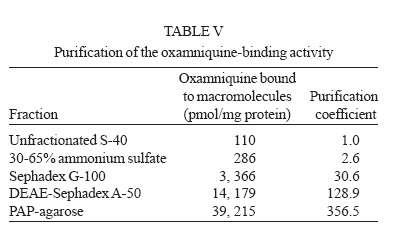
Italy Received 25 May 2006 Code Number: oc06206 Available evidence suggests that the antischistosomal drug oxamniquine is converted to a reactive ester by a schistosome enzyme that is missing in drug-resistant parasites. This study presents data supporting the idea that the active ester is a sulfate and the activating enzyme is a sulfotransferase. Evidence comes from the fact that the parasite extract loses its activating capability upon dialysis, implying the requirement of some dialyzable cofactor. The addition of the sulfate donor 3'-phosphoadenosine 5'-phosphosulfate (PAPS) restored activity of the dialyzate, a strong indication that a sulfotransferase is probably involved. Classical sulfotransferase substrates like b-estradiol and quercetin competitively inhibited the activation of oxamniquine. Furthermore, these substrates could be sulfonated in vitro using an extract of sensitive (but not resistant) schistosomes. Gel filtration analysis showed that the activating factor eluted in a fraction corresponding to a molecular mass of about 32 kDa, which is the average size of typical sulfotransferase subunits. Ion exchange and affinity chromatography confirmed the sulfotransferase nature of the enzyme. Putative sulfotransferases present in schistosome databases are being examined for their possible role as oxamniquine activators. Key words: Schistosoma mansoni - oxamniquine - mechanism of action - sulfotransferase - drug activation Oxamniquine (OXA) has been the cornerstone of Brazilian schistosomiasis control programs during the last 25 years and still is one of the two commercially available drugs that are effective against Schistosoma mansoni (Coura & Amaral 2004, WHO 2002). Available information on the mechanism of action of OXA (and of the related agent hycanthone) suggests that the drug becomes active only upon transformation to an alkylating agent by an enzyme that is present in OXA-sensitive schistosomes (reviewed in Cioli et al. 1995). The evidence originates from genetic crosses between OXA-sensitive and OXA-resistant S. mansoni showing that resistance is a recessive trait, thus implying the existence of a parasite factor that is required for drug activity (Cioli et al. 1992). In addition, a soluble extract from sensitive worms (but not from resistant worms) was capable of converting OXA into an alkylating agent that could bind covalently to the DNA and other macromolecules of the parasite (Pica-Mattoc-cia et al. 1989, 1992). Extracts from S. japonicum or S. haematobium (which are not sensitive to OXA), as well as extracts from mammalian tissues, failed to activate OXA. The OXA activating factor was clearly an enzyme, since it was thermolabile, protease sensitive, inactive at 0°C and required ATP + Mg2+ in a cell-free assay (Pica-Mattoccia et al. 1992). The precise nature of the enzyme, however, was not determined, except for the broad assumption that it might be an esterifying enzyme targeting the hydroxymethyl group that is present in the drug and is essential for its activity (Cioli et al. 1985). This study presents evidence supporting the suggestion that the OXA activating enzyme is a sulfotransferase. Activity could be partially purified, but no sufficient pure product was obtained to characterize the enzyme in molecular terms. MATERIALS AND METHODS Parasites - A Puerto Rican strain of S. mansoni displaying a stable sensitivity to oxamniquine (SEN) was used in most experiments (Cioli 1976). A different Puerto Rican S. mansoni displaying a strong resistance to OXA and hycanthone (BRR) was obtained in 1973 from Dr E Bueding (Johns Hopkins University, Baltimore, MD, US) and subsequently maintained at the Institute of Cell Biology, Rome, Italy, where it was also subjected to drug pressure and selection of survivors (Pica-Mattoccia et al. 1992). Adult schistosomes were obtained by perfusion of mice, 7 weeks or more after infection. Enzyme assays - Worm extracts and fractions thereof were assayed for their OXA-activating capacity as described by Pica-Mattoccia et al. (1992). Briefly, male adult schistosomes were homogenized at 0°C in 0.1M Hepes pH 7.4 containing a mix of protease inhibitors (standard buffer) and the homogenate was centrifuged at 40,000 x g to yield a soluble fraction (S-40). Rat liver extracts were prepared in the same way. Ten µl of a mixture containing 50 mM ATP, 50 mM MgCl2 and 2-8 x 105 cpm of 3H-OXA were then added to 90 µl of the soluble extract and incubated for 2 h at 37°C. Tritiated drug was prepared by reduction of OXA aldehyde with 3H-sodium borohydride, following the procedure described for hycanthone (Pica-Mattoccia et al. 1988). The reaction was stopped by the addition of 10 µl of 3M NaCl and 250 µl of chilled ethanol and kept at -20°C for at least 2 h. The reaction tubes were then centrifuged and the pellet of precipitated macromolecules was dissolved in 400 µl PBS containing 0.25% SDS and 200 µg/ml proteinase K. Samples were left 30 min at 50°C, extracted with two volumes of methylene chloride and the aqueous phase was counted in a liquid scintillation spectrometer. The procedure effectively eliminates free tritiated OXA (soluble in organic solvents), while allowing the estimation of macromolecule-bound radioactivity remaining in the aqueous phase (Pica-Mattoccia et al. 1992). Early experiments of this study were carried out using hycanthone, but it was later shown that OXA and hycanthone have an identical mechanism of action (Cioli et al. 1995), and most of the later experiments were carried out using OXA. When hycanthone was used, this is indicated in the text. The quercetin sulfonation assay was carried out using [35S]PAPS (200 Ci/mmole, New England Nuclear, Boston, MA) as co-substrate, essentially as described by Varin et al. (1987). The conversion of b-estradiol to b-estradiol sulfate was monitored using [3H]17b-estradiol (85 Ci/mmol, New England Nuclear, Boston, MA), as described by Oeda et al. (1992). Fractionation of extract - Five ml (about 50 mg protein) of the soluble worm extract (S-40) was fractionated with ammonium sulfate and the material precipitating at 30-65% saturation (preliminarily shown to contain activity) was dissolved in 0.4 ml standard buffer and applied to a Sephadex G-100 column (1.6x100 cm) equilibrated with the same buffer. Elution was carried out at 4°C and the optical density of collected fractions (1 ml) was read at 280 nm to determine the protein profile. Ninety µl of each fraction was then assayed for OXA-macromolecule binding activity as detailed above. Since gel filtration removes small molecules that may be required as cofactors in the reaction, an aliquot of an extract from resistant worms was added to each assay. The column was calibrated using bovine serum albumin (67 kDa), bovine carbonic anhydrase (29 kDa) and cytochrome C (13 kDa). Fractions containing the peak of activity were pooled and then applied to a DEAE-Sephadex A-50 column (0.5 ´ 2 cm). Elution was carried out with a linear gradient (20 ml) of NaCl from 0 to 0.4M in standard buffer and aliquots of fractions were assayed again for activity. Active fractions were dialyzed against standard buffer and applied to a 0.3 ´ 1.2cm column of PAP-agarose (Sigma). Elution was carried out using 8 ml of a linear gradient of PAPS from 0 to 0.4 mM in standard buffer. RESULTS Requirements of the OXA-macromolecule binding reaction -As previously described, soluble extracts from OXA-sensitive schistosomes were able to produce the covalent binding of tritiated drug to DNA and other macromolecules (Archer et al. 1990, Pica-Mattoccia et al. 1992). The reaction required ATP and Mg2+ and did not occur with extracts from OXA-resistant schistosomes. Table I shows that most of the activity was lost if the extract was dialyzed against standard buffer, but the loss was not irreversible, since activity could be restored by the addition of an extract from drug-resistant worms (which is inactive in itself) to the reaction mixture. No reconstitution occurred if the extract from resistant worms was also dialyzed. This implies that a dialyzable component _ in addition to MgATP _ is required for the reaction to occur. Table I also shows that a number of various additions were tested and proved to be ineffective, with the exception of sulfate ions, which were able to restore most of the activity. When inorganic sulfate was replaced by the common sulfate donor in biological systems, 3'-phos-phoadenosine 5'-phosphosulfate (PAPS) (Robbins & Lipmann 1957) a complete reconstitution was produced. Reconstitution occurred even in the absence of ATP addition, indicating that ATP is needed mainly for PAPS formation (Robbins & Lipmann 1958). In contrast, the non-sulfated PAPS analog (3'-phosphoadenosine 5'-phosphate, PAP) not only failed to restore activity, but also was able to inhibit the non-dialyzed extract. This evidence strongly suggests that the enzyme is a sulfotransferase, presumably esterifying OXA by the addition of a sulfate at the hydroxymethyl group that is present in the drug (Cioli et al. 1985). Incidentally, it is worth noting that PAPS could not be substituted by p-nitrophenyl sulfate, the sulfate donor for a special class of bacterial sulfotrans-ferases (Kobashi et al. 1986). OXA resistance is not due to the lack of PAPS - The notion that the sulfate donor PAPS is necessary for the activation of OXA raised the possibility that OXA resistance might be due to the lack of PAPS, or of PAPS-forming enzymes, in resistant worms. To test this hypothesis, PAPS was added to the extract of resistant worms, but this did not produce any covalent binding of tritiated drug to macromolecules (last line of Table I), thus disproving the hypothesis. Inhibitors of the OXA activation reaction - In order to further test the hypothesis that OXA activation consists in a sulfonation reaction, a number of different compounds were tested for their ability to inhibit the drug-binding assay. The first part of Table II lists several compounds that might function as sulfotransferase substrates and might thus exert a competitive inhibition on the reaction. Of all the compounds tested, quercetin and b-estradiol two natural substrates of sulfotransferase-mediated reactions were clearly able to exert a strong inhibition of 3H-OXA binding to macromolecules. Lower levels of inhibition were also exerted by estrone and dehydroepiandrosterone, while insect steroids and some plant polyphenolic substances showed only marginal effects. The second part of Table II lists some compounds that are known to exert various degrees of inhibition on sulfotransferases. Although fairly effective, none of them produced a complete inhibition at the concentrations tested. Quercetin and b-estradiol are sulfonation substrates - Additional evidence for the sulfotransferase nature of the schistosome enzyme was provided by the finding that quercetin and b-estradiol not only were able to compete with OXA in the reaction, but were also able to function as sulfate acceptors under the action of schistosome extracts. Most important, sulfate transfer to the two acceptors was mediated only by extracts of OXA-sensitive schistosomes, while the extracts of drug-resistant worms failed to produce any effect. Table III shows that quercetin could be sulfonated by rat liver extract and that this reaction was not sensitive to competition with the oxamniquine congener, hycanthone. This indicates that the mammalian enzyme does not recognize the antischistosomal drug, as previously demonstrated (Pica-Mattoccia et al. 1992). An extract from sensitive worms was also capable to sulfonate quercetin and this reaction could be competed by free hycanthone, stressing the drug specificity of the parasite enzyme. Only marginal quercetin sulfonation was mediated by an extract from OXA-insensitive schistosomes. Table IV shows that b-estradiol could be easily sulfonated by rat liver enzymes, as expected (Borthwick et al. 1993). An extract from sensitive worms was also capable to produce b-estradiol sulfonation, whereas an extract from OXA-insensitive schistosomes did not produce b-estradiol sulfate above detection level. The amount of sulfate bound to b-estradiol by schistosome extracts was much less than the amount bound under the activity of rat liver extract. This is not completely unexpected when considering that the specificity of the parasite enzyme is directed towards an unknown substrate that is likely to be at best only partially resembling the mammalian steroid substrate. Fractionation and purification of the activating enzyme - An ammonium sulfate fraction of the soluble extract (S-40) of OXA-sensitive schistosomes was subjected to gel filtration analysis under non-denaturing conditions and the fractions were tested for protein content and for 3H-OXA binding activity. Fig. 1 shows that, while most of the protein was eluted with the column front, the bulk of enzymatic activity was eluted in a region corresponding to a molecular weight of about 30-32 kDa, in accordance with the fact that most cytosolic sulfotransferase subunits consist of around 300 amino acid residues (Pe-trotchenko et al. 2001). A pool of the Sephadex fractions containing the peak of activity was applied to a DEAE-Sephadex column and eluted with a NaCl gradient. Fig. 2A shows that activity eluted as a single peak at about 0.2M salt concentration, in agreement with the behavior of other sulfotransferases. Fractions from DEAE-Sephadex containing activity were pooled, dialyzed against standard buffer and applied to a PAP-agarose affinity column. Elution was performed with a gradient of PAPS and the results are shown in Fig. 2B. The single peak of activity eluting with the specific co-substrate adds further weight to the evidence that the enzyme is indeed a sulfotransferase. As it happens with many sulfotransferases, the OXA-macromolecule binding activity proved to be rather labile, since it was gradually lost upon simple storage at 4°C, at the rate of about 20%/week. This makes it difficult to keep an accurate bookkeeping of activity through the various steps of the purification procedure. Nevertheless, an approximate probably underestimated accounting of the purification achieved is given in Table V. It should be noted that even the most purified fraction obtained from PAP-agarose still presented several bands when analyzed by SDS-PAGE (not shown). DISCUSSION This study contributes additional evidence to the available information regarding the proposed mechanism of action of OXA (Cioli et al. 1995). It was previously shown that an enzymatic activity present in OXA-sensitive schistosomes (but absent in drug resistant worms) could produce the covalent binding of OXA to the DNA and other macromolecules of the parasite (Pica-Mattoccia et al. 1992). It was postulated that the schistosome enzyme was inducing the formation of an OXA ester that would then spontaneously dissociate giving rise to an electrophilic moiety capable of alkylating parasite macromolecules (Cio-li et al. 1985, 1995). The chemical nature of the postulated ester was, however, unknown and a direct identification was problematic in view of the very short half life that such an unstable compound would exhibit. Data presented here strongly support the notion that the ester is a sulfate and the OXA activating enzyme is a sulfotransferase. The initial observation that a dialyzable cofactor is essential for the activation reaction prompted us to examine a number of potential compounds that could restore activity to the dialyzed schistosome extract. Inorganic sulfate provided a substantial re-activation and the organic sulfate donor PAPS (Robbins & Lipmann 1957) proved capable of full restoration of activity. We thus inferred that the intact parasite extract probably contains a certain amount of preformed PAPS and is very likely to contain the two key enzymes for PAPS formation from ATP and inorganic sulfate, i.e. ATP sulfurylase and adenosine 5'-phosphosulfate kinase (Robbins & Lipmann 1958). Dialysis removes PAPS and its two building blocks (MgATP and inorganic sulfate), thus impeding the reaction to proceed. Addition of inorganic sulfate may restore activity if ATP and the two enzymes are present in adequate amounts, while addition of PAPS is obviously expected to be fully effective. A non-dialyzed extract from drug resistant worms, which is inactive per se, is capable of restoring activity because it can supply PAPS and the whole PAPS-forming machinery. PAPS donates the sulfate and is converted to PAP in the sulfotransferase mediated reaction. PAP proved unable to restore activity to the dialyzed extract and, being a reaction product, actually inhibited the activity of the intact extract. The requirement of PAPS in the reaction of OXA activation raised the theoretical possibility that drug resistant schistosomes might fail to activate the drug because they are deficient in PAPS, but PAPS addition to the extract from resistant worms failed to confer activity, thus disproving the hypothesis. To further validate the notion that OXA activation involves a sulfonation reaction, we asked whether known sulfotransferase substrates were able to compete with OXA in the reaction. Several compounds possessing one or more alcoholic functions the main target of sulfonation reactions were tested and some of them were indeed able to produce a strong competition, like the flavonoid quercetin and some mammalian steroids. The fact that only a restricted number of structures were able to compete indicates that the putative schistosome sulfotransferase, like other members of the sulfotrans-ferase family, could have a rather stringent substrate specificity (Glatt 1997). A few of the known non-competitive inhibitors of sulfotransferases gave variable, but incomplete inhibition of the oxamniquine-macromolecule binding reaction. We interpret this as an indication that the schistosome enzyme has relatively unusual characteristics with respect to canonical sulfotransferases. In addition, at least two of the competing compounds, i.e. quercetin and b-estradiol, were able to function as sulfate acceptor substrates under the action of a schistosome extract, when tested in classical sulfonation assays based on the use of 35PAPS and radioactive b-estradiol, respectively. The fact that extracts from OXA resistant worms were unable to produce the sulfonation of these heterologous substrates is strong evidence to support the idea that this type of drug resistance is indeed the result of a missing sulfonation activity. By the same argument, OXA activity must require a sulfonation reaction. Ancillary evidence supporting the assumption that the OXA activating enzyme is a sulfotransferase, comes from the biochemical characteristics of the schistosome-derived activity. The schistosome enzyme is found in the cytosol and elutes in gel filtration in a region corresponding to about 32 kDa under non-denaturing conditions. Most mammalian and plant cytosolic sulfotransferases exhibit a native size that is twice this value and have been shown to be dimers consisting of two identical subunits. There are, however, examples of native sulfotransferases that consist of a single subunit, as is the case when a single valine to glutamic acid substitution occurs in a dimerization motif near the C-terminus of the protein (Petrotchenko et al. 2001). The schistosome enzyme apparently belongs to this subset of monomeric proteins. Ion exchange chromatography has often been used for the isolation of sulfotransferases (Falany et al. 1990) and the schistosome activity proved also amenable to this type of purification. The peak of activity was eluted from DEAE-Sephadex at the NaCl concentration of 0.2 M, but such a peak still consisted of about 20 different proteins when analyzed by SDS-PAGE. In a further purification step, the DEAE-Sephadex peak was applied to a PAP-agarose affinity chromatography support and proved capable of binding and of subsequent elution with the specific sulfotransferase substrate PAPS. This constitutes additional evidence for the sulfotransferase nature of the enzyme. Although each purification step provided some increase in the level of specific activity, no electrophoretically pure fraction could be obtained. In addition, activity proved to be quite labile and was gradually lost even upon simple storage at 4°C, as it occurs with other sulfotrans-ferases. Thus, we were unable to obtain sufficient amounts of high purity protein for further structural studies. A number of heterologous anti-sulfotransferase antibodies were tested, but none of them gave consistent results when assayed in Western blots against schistosome extracts. This prevented us from attempting to clone the gene by library immunoscreening. Also, we were not successful with library screening using degenerate primers based on sulfotransferase sequences known in other species. The recent availability of extensive S. mansoni EST databases and the progress in the schistosome genome sequencing (Verjovski-Almeida et al. 2003, LoVerde et al. 2004), opens the possibility of an in silico analysis of schistosome sulfotransferases, including the putative OXA-activating enzyme described in this study. Work is in progress along these lines. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06206f2.jpg] [oc06206t4.jpg] [oc06206t1.jpg] [oc06206t5.jpg] [oc06206f1.jpg] [oc06206t3.jpg] [oc06206t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}