 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
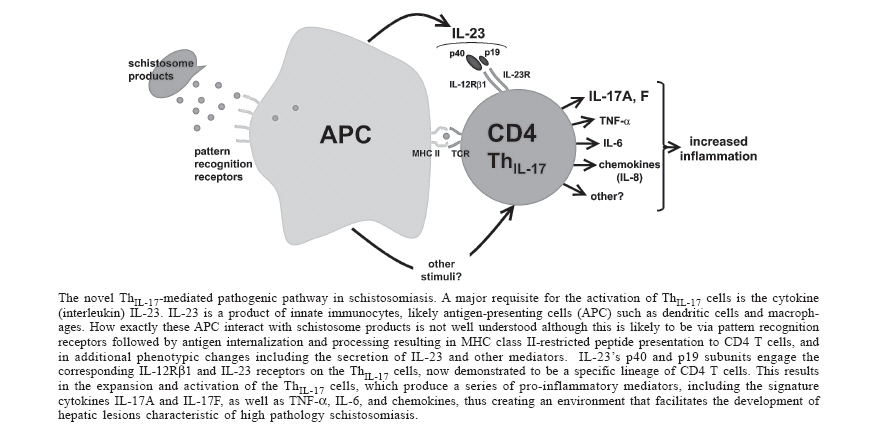
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101,Suppl. I, October ,2006, pp. 327-330 CD4 T cells producing pro-inflammatory interleukin-17 mediate high pathology in schistosomiasis Laura I Rutitzky, Miguel J Stadecker+ Department of Pathology, Tufts University School of Medicine,
150 Harrison Ave, Boston, MA 02111, US
Financial support: Public Health Service grants RO1-18919 and RO1-48736 Received 25 May 2006 Code Number: oc06210 In murine schistosomiasis mansoni, pronounced CD4 T cell-mediated, egg-induced, hepato-intestinal immunopathology and death, whether genetically determined or elicited experimentally, are associated with failure to down-regulate a net pro-inflammatory immune response. Important evidence contributing to this notion comes from the observation that immunization with schistosome egg antigens in CFA (SEA/CFA) causes low pathology C57BL/6 mice to develop an exacerbated form of disease and death in a cytokine milieu characterized by elevated interferon (IFN)-g levels. Since such a pro-inflammatory environment presumes a signaling pathway involving interleukin (IL)-12, the SEA/CFA immunization model was used to examine the extent of hepatic immunopathology in the absence of this cytokine. Surprisingly, the IL-12p40 subunit was an absolute requirement for the development of exacerbated disease, whereas the IL-12p35 subunit was not. Moreover, significantly elevated in vitro production of IL-17, but not of IFN-g, correlated with the high pathology, and neutralization of IL-17 in vivo resulted in a significant reduction of hepatic inflammation. Our findings clearly demonstrate the pathogenic potential of the novel IL-17-producing T cell subpopulation (ThIL-17), previously shown to mediate chronic inflammation in autoimmune disease. They also imply that IL-23, but not IL-12, is the critical signal necessary to support the pro-inflammatory ThIL-17 subset involved in high pathology schistosomiasis. Key words: schistosomiasis - severe immunopathology - CD4 T cells - interleukin-17 In schistosomiasis caused by the helminth trematode species Schistosoma mansoni, the hepato-intestinal gra-nulomatous and fibrosing inflammation precipitated by parasite eggs is mediated by CD4 T cells. Human patient populations infected with this parasite display a great variation in the magnitude of clinical disease. Likewise, in the murine model, the CBA and C3H strains typically develop a more pronounced hepatic immunopathology characterized by larger egg granulomas with hepatic parenchymal inflammation, whereas in C57BL/6 (BL/6) mice the lesions are uniformly less severe (Cheever et al. 1987, Rutitzky et al. 2005a). Several independent lines of evidence have linked the high pathology to the persistence of a net pro-inflammatory state marked by the increase of Th1-type cytokines such as interferon (IFN)-g and tumor necrosis factor (TNF)-a, the deficiency of anti-inflammatory Th2-type cytokines such as interleukin (IL)-4 and IL-10, or both. For example, schistosome egg antigen (SEA)-stimulated mesenteric lymph node cells from 7 week-infected high pathology CBA mice produce significant amounts of IFN-g, whereas those from low pathology BL/6 mice do not (Rutitzky et al. 2001, 2003a, 2005a). Moreover, mice with impaired costimulatory B7-CD28 (King et al. 1996, Hernandez et al. 1999), CD40-CD154 (MacDonald et al. 2002) and B7RP-1-ICOS (Rutitzky et al. 2003b) systems, or deficient in anti-inflammatory cytokines IL-4 (Brunet et al. 1997) and IL-10 (Hoffmann et al. 2000, Sadler et al. 2003), or in B cells (Hernandez et al. 1997) or in alternatively activated macrophages (Herbert et al. 2004), display a pro-inflammatory cytokine dominance and are prone to pathology exacerbation. Lastly, immunization with SEA in CFA causes the typically low pathology C57BL/6 mice to develop an exacerbated form of disease and death in a cytokine milieu characterized by elevated IFN-g levels (Rutitzky et al. 2001). In essence, a net pro-inflammatory host immune reactivity to egg antigens results in enhanced pathology that manifests itself early after patent infection and often is associated with premature death. The development of Th1 vs Th2 CD4 cell subtypes endowed with contrasting inflammatory capabilities is the product of distinct activation programs operating in common undifferentiated precursor cells. The signal transducer and activation of transcription factors (STAT) 1 and 4, the transcription factor T-bet and IL-12 are involved in Th1 differentiation (Szabo et al. 2003), whereas STAT 6, GATA-3 and IL-4, respectively, facilitate Th2 development (Zhu et al. 2006). The work presented here and at the 10th International Symposium on Schistosomiasis, held in Belo Horizonte, Brazil, on September 25-28, 2005, focused specifically on the role of IL-12 in promoting the pro-inflammatory state associated with severe egg-induced immunopathology. IL-12 is a heterodimeric 70 kDa cytokine composed of two subunits of 40 and 35 kDa, termed IL-12p40 and IL-12p35 (Trinchieri 1998, 2003a, Trinchieri et al. 2003). IL-12 is produced by activated innate immunocytes, such as dendritic cells (DC), and is recognized by lymphocytes expressing IL-12Rb1 and IL-12Rb2 receptors for the IL-12p40 and IL-12p35 subunits, respectively, thereby inducing IFN-g and other pro-inflammatory mediator production (Trinchieri 1998, 2003). Effective examination of the role of IL-12 in the development of schistosome immunopathology requires the use of mice deficient in either of its subunits, IL-12p40 (IL-12p40-/-) and IL-12p35 (IL-12p35-/-). Because such mice are currently only available on a genetic background (BL/6) conducive to low egg-induced pathology, they were concomitantly immunized with SEA in CFA (Rutitzky et al. 2001). This treatment has been demonstrated to cause in infected BL/6 mice a marked enhancement of the hepatic lesions and early death in a pro-inflammatory environment, a situation comparable to that seen in other mouse strains which are naturally prone to high pathology. Schistosome-infected IL-12p40-/- mice immunized with SEA/CFA indeed displayed markedly reduced hepatic pathology, which was no different from that seen in unimmunized controls. However, in contrast, SEA/CFA-immunized IL-12p35-/- mice exhibited the accelerated death and exacerbated hepatic lesions typically seen in similarly treated BL/6 WT mice (Rutitzky et al. 2005b). Notably, the dramatic dichotomy in disease severity between the IL-12p40-/- and IL-12p35-/- mice was evident despite the fact that both groups had extremely low levels of pro-inflammatory IFN-g (Rutitzky et al. 2005b). These findings clearly demonstrated that only IL-12p40 was essential for the development of high pathology whereas IL-12p35 (and thus the dimeric form IL-12p70) were not, and, additionally, that there was no correlation between the severity of immunopathology and the levels of IFN-g (Rutitzky et al. 2005b). Surprisingly, the high pathology state most closely correlated with elevated levels of IL-17 produced by SEA-stimulated granuloma and mesenteric lymph node T cells (Rutitzky et al. 2005b). A compelling reason for investigating IL-17 was because this cytokine has recently received considerable attention as a novel pro-inflammatory mediator of chronic inflammation associated with cell-mediated autoimmune disease. Two models in which IL-17 has been clearly shown to play a decisive role in pathogenesis are experimental allergic encephalomyelitis (EAE) (Cua et al. 2003, Langrish et al. 2005) and collagen induced arthritis (CIA) (Murphy et al. 2003). In both EAE and CIA, the IL-17 was produced by a population of CD4 T cells expressing the CD69 activation marker and a CD44high, CD62low memory phenotype (Aggarwal et al. 2003, Langrish et al. 2005). In our system, cells with these characteristics were similarly increased in the large egg granulomas from the SEA/CFA-immunized WT and IL-12p35-/-, but not in those from the IL-12p40-/- mice. Moreover, neutralization of IL-17 did result in an inhibition of immunopathology, denoting a direct role for this cytokine in the pathogenesis of the enhanced granulomatous inflammation. Importantly, elevated levels of IL-17 were also detected in schistosome-infected CBA mice, which are naturally prone to high pathology, and their lesions were similarly reduced following treatment with neutralizing anti-IL-17 antibodies (Rutitzky et al. 2005b). These findings clearly tie the ThIL-17 cells to the development of high-pathology schistosomiasis. IL-17A and IL-17F are the members of the IL-17 cytokine family responsible for the pathogenic activity of the ThIL-17 cells, as the antibodies resulting in disease amelioration are directed against these species (Lubberts et al. 2004, Rutitzky et al. 2005b). However, other pro-inflammatory mediators produced by the ThIL-17 cells, including TNF-a, IL-6, IL-8 and other chemokines (Kolls & Linden 2004, Langrish et al. 2004), may also play a significant role in the pathology. One apparent mechanism used by ThIL-17 cells to cause pathology is through recruitment of granulocytes, which in the case of the schistosome egg-granulomas, may include the eosinophils. This issue is currently under investigation. Current models about the development of ThIL-17 cells place this subset directly under the control of IL-23, but not of IL-12 (Trinchieri et al. 2003, McKenzie et al. 2006). IL-23 is a member of the IL-12 family of heterodimeric cytokines which utilizes the same IL-12p40 subunit in association with a distinct p19 subunit (Oppmann et al. 2000). It thus becomes evident that it is the absence of IL-23 formation in SEA/CFA-immunized, schistosome-infected mice lacking the IL-12p40 subunit that explains their low levels of immunopathology and of IL-17. Like IL-12, IL-23 is a product of stimulated innate immunocytes such as DC and macrophages, which activate T cells bearing the IL-12Rb1 receptor for the common IL-12p40 subunit and a specific IL-23 receptor for the p19 subunit (Parham et al. 2002). Unlike IL-12, the binding of IL-23 to its receptors induces phosphorylation of STAT 3 in addition to STAT 4 (Trinchieri et al. 2003, Holscher 2004), with further differences in signal transduction to be elucidated. It has been recently demonstrated that the ThIL-17 cells activated by IL-23 constitute a fully distinct, de novo, pro-inflammatory T cell differentiation line, which does not branch off from pro-inflammatory Th1 cells activated by IL-12, as first surmised to be the case (Harrington et al. 2005, Park et al. 2005). The Figure illustrates the likely activation pathway and function of the CD4 ThIL-17 cell subset in infection with schistosomiasis. While there is ample evidence that these cells are stimulated by IL-23 and mediate severe immunopathology, multiple questions regarding the pathogenic IL-23/ThIL-17 pathway still remain to be answered. For example, it is not clear if and how schistosomes, particularly the highly immunopathogenic eggs, directly activate innate immunocytes or other potential cell sources of IL-23. A subject of considerable discussion is about these cells possessing dedicated receptors that recognize specific molecular patterns from schistosomes equivalent to the toll-like receptors (TLR), C-type lectins and other molecules that recognize microbial motifs (discussed in Hokke & Yazdanbakhsh 2005). Other questions concern the variation in IL-23 production by different innate immunocytes and the nature of possible additional mediators capable of stimulating ThIL-17 cells; this issue can be addressed by examination of schistosome-infected mice lacking the IL-23-specific p19 subunit. Finally, regarding the ThIL-17 cells themselves, it is not clear whether they all are of the CD4 type; moreover, their kinetics of IL-23R expression, precise costimulation requirements and full range of secreted pro-inflammatory mediators remain to be fully elucidated. Conclusions Research into the basis of the dissimilar immunopathology development in murine schistosomiasis has revealed a critical pathogenic role of a novel pro-inflammatory CD4 T cell subset producing IL-17 (ThIL-17), which is specifically detected in situations of high pathology, either induced or genetically determined. Clearly, the ThIL-17 cells are stimulated by the heterodimeric cytokine IL-23, for which they possess specific receptors. The origin of IL-23 and other possible activating signals is ascribed to innate immunocytes, although the precise pathogen-derived stimuli are not known. ThIL-17 cells have been shown to participate in an increasing number of host responses stimulated by a variety of self antigens and pathogens (Infante-Duarte et al. 2000, Cua et al. 2003, Murphy et al. 2003, Lubberts et al. 2004, Happel et al. 2005, Khader et al. 2005, Langrish et al. 2005, Kleinschek et al. 2006, Mensah-Brown et al. 2006). It now is of great interest and importance to ascertain their presence and activity in human diseases including schistosomiasis. References
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06210f1.jpg] |
| |||||||||

{kind=link}