 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 101,Suppl. I, October ,2006, pp. 365-368 Aluminum hydroxide associated to Schistosoma mansoni 22.6 kDa protein abrogates partial protection against experimental infection but not alter interleukin-10 production Lucila GG Pacífico, Cristina T Fonseca, Michele M Barsante, Luciana S Cardoso*, Maria Ilma Araújo*, Sérgio C Oliveira/+ Departamento de Bioquímica e Imunologia e Instituto
de Investigação em Imunologia, Instituto Millennium, Universidade
Federal de Minas Gerais, Av. Antônio Carlos 6627, 31270-901 Belo Horizonte,
MG, Brasil *Serviço de Imunologia, Hospital Universitário Professor
Edgard Santos, Salvador, BA, Brasil
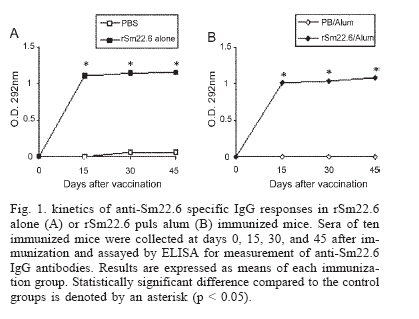
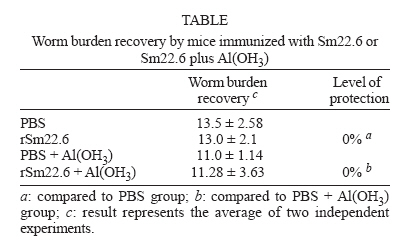
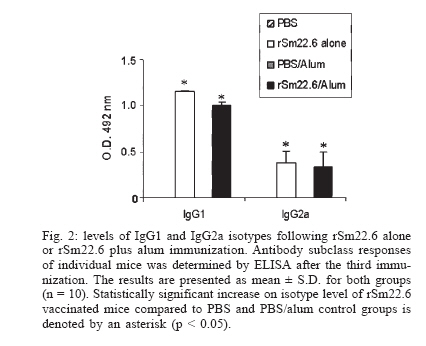
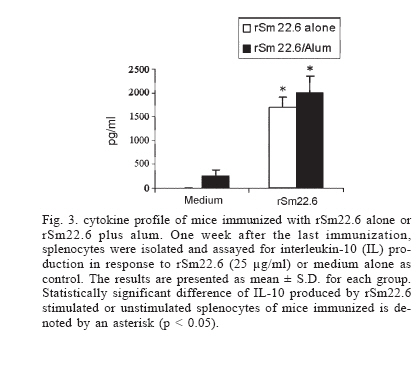
Financial support: CNPq, PADCT/CNPq, Fapemig, Fapesp Received 25 May 2006 Code Number: oc06218 The need to develop a vaccine against schistosomiasis led several researches and our group to investigate proteins from Schistosoma mansoni as vaccine candidates. Sm22.6 is a protein from S. mansoni that shows high identity with Sj22.6 and Sh22.6 (79 and 91%, respectively). These proteins are associated with high levels of IgE and protection to reinfection. Previously, we have shown that Sm22.6 induced a partial protection of 34.5% when used together with Freund's adjuvant and produced a Th0 type of immune response with interferon-g and interleukin-4. In this work, mice were immunized with Sm22.6 alone or with aluminum hydroxide adjuvant and high levels of IgG, IgG1, and IgG2a were measured. Unfortunately, no protection was detected. Since IL-10 is a modulating cytokine in schistosomiasis, we also observed a high level of this molecule in splenocytes of vaccinated mice. In conclusion, we did not observe the adjuvant effect of aluminum hydroxide associated with rSm22.6 in protective immunity. Key words: Schistosoma mansoni - Sm22.6 - recombinant vaccine - aluminum hydroxide - adjuvant - interleukin-10 In many development countries Schistosoma mansoni is the most widespread species causative of schistosomiasis. Parasite eggs laid in the hepatic portal vasculature are the principal cause of the morbidity since they cause a granulomatosous inflammation when retained in the liver and severe pathology may be fatal (King 2001). Schistosomiasis has been treated by chemotherapy using praziquantel. However, drug resistant strains have been described which reinforce the necessity to develop a vaccine against schistosomiasis (Doenhoff et al. 2002). Several investigators have focused on protective immune against schistosomiasis using subunit vaccine protocols, but until now none of them was able to induce more than 67% (Zouain et al. 2001) of protection. S. mansoni have a complex life cycle and sexual dimorphism and this turns difficult to find a target protein to be used as an effective vaccine. Nevertheless the successful vaccination of both rodents and primates with attenuated cercariae indicates that vaccine development is possible (Coulson 1997). The S. mansoni genome project has identified several new genes coding for proteins that have been studied as candidates to vaccine (Verjovski-Almeida et al. 2003). In our laboratory some proteins have been studied such as Sm14 (Fonseca et al. 2004), Sm29 (Cardoso et al. 2006), paramyosin (Fonseca et al. 2005), and Sm22.6 (Pacífico et al. 2006) in the field of vaccine development. Sm22.6 is a protein of 22.6 kDa with high similarity to Sj22.6 (S. japonicum) and Sh22.6 (S. haematobium) which are the main targets for the human IgE response, and high levels of IgE are correlated with resistance to re-infection after chemotherapy (Dunne et al. 1997, Santiago et al. 1998). Sm22.6 is a protein that showed partial protection of 34.5% when immunized using Freund's adjuvant (CFA) showing high levels of IgG1, production of interferon-g and interleukin-4 resulting in a mix of Th1 and Th2 response (Pacífico et al. 2006). CFA continues to be the most commonly used in mice despite their potential hazards and it is not recommended for use in human. Aluminum salts are generally weaker adjuvants but they are the primary adjuvants used in humans due to safety, cause mild inflammatory reactions and are efficient for generating memory. So next, we have tested the rSm22.6 alone and rSm22.6 with aluminum hydroxide Al(OH)3 as adjuvant and although high levels of IgG, IgG1, and IgG2a were present, rSm22.6 alone or with Al(OH)3 did not induce protection against challenge infection. MATERIALS AND METHODS Mice and parasites - C57BL/6 female, 6-8 weeks old, were obtained from the Federal University of Minas Gerais (UFMG) animal facility. Cercariae of S. mansoni (LE strain) was maintained routinely on Biomphalaria glabrata snails at GIDE (Grupo Interdisciplinar de Estudo da Esquistossomose)-UFMG and prepared by exposing infected snails to light for 1 h to induce shedding. Cercarial numbers and viability were determined using a light microscope prior to infection. Sm22.6 recombinant protein - The rSm22.6 was produced as previously described by our group (Pacífico et al. 2006). Briefly, expression of the fusion protein composed by Sm22.6 and maltose-binding protein (MBP) was induced by 0.6mM of IPTG (isopropyl-B-D-galac-topyranoside). After 3 h at 37°C and 180 rpm, the bacterial cells were harvested by centrifugation at 4000 ´ g for 20 min. The pellet was ressuspended in 100 ml of phosphate buffer saline (PBS, pH 8.4) containing 25 mg of lysosyme and then suffered heat and cold lyse and was submitted to three cycles of 30 s sonication to release the fusion protein, which was purified through affinity chromatography using amylose resin. Mice immunization - Mice were divided into four groups of ten mice each. C57BL/6 mice were subcutaneously (s.c.) injected in the nape of the neck with 25 µg of rSm22.6 on days 0, 15, and 30. The recombinant protein was used in mice immunization alone or with aluminum hydroxide adjuvant (Alum). In the control groups, PBS alone or alum in PBS was administered using the same im-munization protocol. Challenge infection and worm burden recovery - Fifteen days after the last boost, mice were challenged through percutaneous exposure of abdomen skin for 1 h in water containing 30 cercariae (LE strain). Forty-five days after challenge, adult worms were perfused by portal veins (Smithers & Terry 1965). Protection was calculated comparing the number of worm recovered from each group of immunization with their respective controls, using the formula below: Burden recovered from control group - Measurement of specific anti-Sm22.6 antibodies - Following immunization, sera of ten mice from each vaccinated or control groups were collected at 2-week interval. The measurement of specific anti-Sm22.6 antibodies was performed using indirect ELISA. Maxisorp 96-well microtiters plates (Nunc, Denmark) were coated with 5 µg/ml of rSm22.6 in carbonate-bicarbonate buffer, pH 9.6 for 12-16 h at 4°C, then blocked for 2 h at room temperature with 200 µl/well of PBST (phosphate buffer saline, pH 7.2 with 0.05% Tween-20) plus 10% FBS (fetal bovine sera). A hundred microliters of each sera diluted 1:100 in PBST was added per well and incubated for 1 h at room temperature. Plate-bound antibody was detected by peroxidase-conjugated anti-mouse IgG (Promega) diluted in PBST 1:10,000, and IgG1 (Sigma) and IgG2a (Sigma) diluted in PBST 1:5000 and 1:2000, respectively. Color reaction was developed by addition of 100 µl/well of 200 pmoles of OPD (o-phenylenediamine, Sigma) in citrate phosphate buffer, pH 5.0 plus 0.04% H2O2 for 10 min and stopped with 50 µl of 5% sulfuric acid per well. The plates were read at 492 nm in an ELISA reader (BioRad, Hercules, CA). Cytokine analysis - Splenocytes were isolated from macerated spleen of individual mice one week after the third immunization with rSm22.6 alone or rSm22.6 plus alum and washed twice with sterile PBS. After washed, the cells were adjusted to 1 ´ 106 cells per well for IL-10 assay in RPMI 1640 medium (Gibco) supplemented with 10% FBS, 100 U/ml of penicillin G sodium, 100 mg/ml of streptomycin sulfate, 250 ng/ml of amphotericin B. Splenocytes were maintained in culture with medium alone or stimulated with rSm22.6 (25 µg/ml). The 96-well plates (Nunc) were maintained in an incubator at 37ºC with 5% of CO2. Culture supernatants were collected after 72 h of rSm22.6 stimulation for cytokine analysis. The assay for measurement of IL-10 was performed using the BD OptEIA ELISA kit (BD Bioscience Pharmigen, San Diego, CA) according to manufacture's directions. Statistical analysis - Statistical analysis was performed with Student's t test using a computer software package GraphPad Prism4 (GraphPad Software, San Diego, CA). RESULTS IgG responses to rSm22.6 following immunization - To investigate the presence of specific anti-Sm22.6 IgG antibodies, sera of ten vaccinated animals of each group were tested by ELISA. rSm22.6 alone or plus alum vaccinated group showed high titers of IgG that appeared 15 days after the first immunization compared to control group (Fig. 1). The presence of aluminum hydroxide adjuvant did not enhance the levels of rSm22.6 specific IgG, when compared to the rSm22.6 alone vaccinated group. rSm22.6 alone or plus alum did not confer protective immunity - To measure the protection level induced by rSm22.6 alone or plus alum immunization, mice were challenged and the level of protection was determined by the difference in the number of adult worm recovered in the experimental group compared to each control group 45 days post-challenge. Mice immunized with rSm22.6 alone or plus alum showed no protection against experimental infection when compared to control group (PBS or PBS + Al(OH)3 ) (Table). IgG1/IgG2a production after rSm22.6 vaccination - We investigate which type of immune response is induced by vaccination with rSm22.6 alone and rSm22.6 plus alum. The detection of IgG1 as a Th2 marker and IgG2a as a Th1 marker was performed by ELISA. Both treatments showed the same levels of IgG1 and IgG2 and statistically significant compared to control group (Fig. 2). rSm22.6 alone or plus alum immunization induces IL-10 production - IL-10 is a regulatory and anti-inflammatory cytokine and it is important to modulate the immune response. At seven days after the third dose of vaccine significant levels of IL-10 was detected in the supernatant of splenocyte culture of mice immunized with rSm22.6 alone or rSm22.6 plus alum when these cells were stimulated with rSm22.6 (Fig. 3). DISCUSSION Schistosomiasis is a tropical disease that affects more than 200 million people worldwide (Chitsulo et al. 2000). Although schistosomiasis is effectively treated with praziquantel, rapid reinfection with rebound morbidity precludes effective control based on chemotherapy alone and justifies current efforts to develop vaccines against this parasite. Antigens from S. mansoni have been tested as vaccine to immunized mice against challenge with cercariae. One of them, Sm22.6 is a protein with 22.6 kDa that is found in all life cycle except egg (Stein & David 1988) and it has 91% of identity to Sh22.6 (S. haematobium) and 79% to Sj22.6 (S. japonicum). IgE from patients resistant to re-infection after treatment recognizes rSm22.6 indicating that Sm22.6 induces a protective response against S. mansoni (Webster et al. 1996). rSm22.6 has been used in a vaccine model plus CFA and induced 34.5% protection (Pacífico et al. 2006). Even though CFA is considered to be one of the most effective adjuvants, there are problems and hazards associated with its use (Steiner et al. 1960, Broderson 1989, Claassen et al. 1992, Kleinman et al. 1993). In this study, we evaluated the protective efficacy of the rSm22.6 alone or rSm22.6 plus Alum. Immunization with rSm22.6 alone or rSm22.6/Alum successfully mounted a specific anti-Sm22.6 IgG antibody response in C57BL/6 mice, however IgG levels did not increase by the presence of alum. Regarding isotypes, C57BL/6 mice from both groups immunized with rSm22.6 alone or rSm22.6/Alum produced similar same levels of IgG1 and IgG2a anti-Sm22.6 when compared with their control groups. It was surprising since alum is an adjuvant that drives Th2 responses and we did not observe increase in anti-Sm22.6 IgG1 or decrease in IgG2a isotype in alum vaccinated mice. To determine if rSm22.6 alone or rSm22.6/Alum conferred protection against S. mansoni infection, both groups were challenged with two weeks after the last boost and 45 days later worm burdens were analyzed. Immunization with rSm22.6 or rSm22.6/alum did not induce protection against cercarial challenge. There was no apparent correlation between the antibody titers generated and protective efficacy, since all immunized groups had high levels of antibody. The explanation may be that only production of antibodies to this particular antigen is insufficient to induce protection against S. mansoni infection. Jankovic et al. (1999) has demonstrated using B cell knockout mice that an effective vaccination against schistosomes depends on the simultaneous induction of both humoral and cell-mediated immunity. Additionally, data from double-cytokine knockout mice suggest that the establishment of a robust cellular and humoral response is probably the key for generating maximal immunity to schistosomes (Wynn & Hoffmann 2000). Another point could be the fact that rSm22.6 alone or rSm22.6 plus alum immunization induced high levels of IL-10 production. IL-10 is a regulatory cytokine and its production modulates the inflammatory response that is important to protect mice against murine schistosomiasis. ACKNOWLEDGMENTS To Florence Mara Rosa, for technical assistance in challenge infection, and Aparecida Soares for mice perfusion. To the Laboratory of Helminthes Immunology of the Department of Parasitology at UFMG for providing S. mansoni cercariae for challenge infection experiments. REFERENCES
Copyright 2006 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc06218f1.jpg] [oc06218f3.jpg] [oc06218f2.jpg] [oc06218t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}