 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
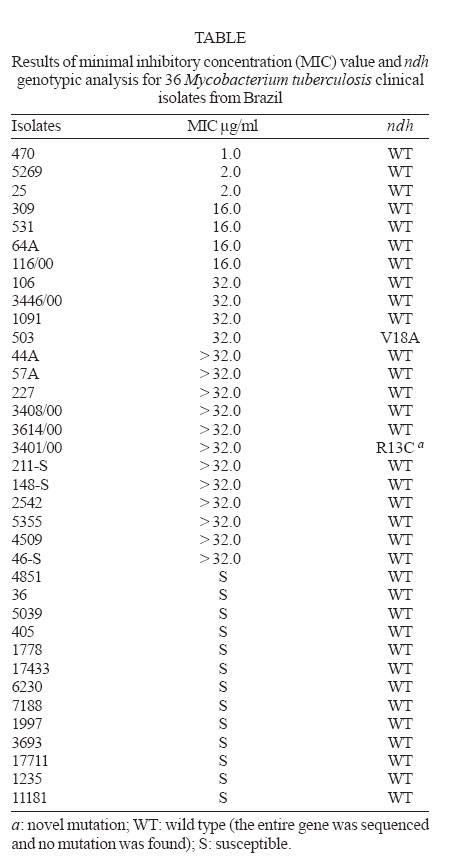
Memórias do Instituto Oswaldo Cruz, Vol. 102, No.1, February 2007, pp. 59-61 Characterization of ndh gene of isoniazid resistant and susceptible Mycobacterium tuberculosis isolates from Brazil Rosilene Fressatti Cardoso/+, Marco Antonio Cardoso, Clarice Queico Fujimura Leite*, Daisy Nakamura Sato**, Elsa Masae Mamizuka***, Rosário Dominguez Crespo Hirata***, Fernando Fiúza de Mello****, Mario Hiroyuki Hirata*** Laborartório
de Bacteriologia Clínica, Departamento de Análises Clínicas,
Universidade Estadual de Maringá, Av. Colombo 5790, Bloco J-90, Jardim
Universitário, 87020-900 Maringá, PR, Brasil *Departamento
de Ciências Biológicas, Unesp, Araraquara, SP, Brasil **Instituto
Adolfo Lutz, Ribeirão Preto, SP, Brasil ***Universidade de São
Paulo, São Paulo, SP, Brasil ****Instituto Clemente Ferreira, São
Paulo, SP, Brasil Financial support: Fapesp, Capes Received 6 July
2006 Code Number: oc07009 Resistance in Mycobacterium tuberculosis to isoniazid (INH) is caused by mutations in the catalase-peroxidase gene (katG), and within the inhA promoter and/or in structural gene. A small percentage (~ 10%) of INH-resistant strains do not present mutations in both of these loci. Other genes have been associated with INH resistance including the gene encoding for NADH dehydrogenase (ndh). Here we report the detection of two ndh locus mutations (CGT to TGT change in codon 13 and GTG to GCG change in codon 18) by analyzing 23 INH-resistant and in none of 13 susceptible isolates from Brazilian tuberculosis patients. We also detected two isolates without a mutation in ndh, or any of the other INH resistance-associated loci examined, suggesting the existence of additional, as yet to be described, INH resistance mechanisms. Key words: Mycobacterium tuberculosis - NADH dehydrogenase gene - isoniazid resistance Isoniazid (INH) has been a first-line drug for tuberculosis treatment for many years and yet its mechanism of action remains only partially understood (Rozwarski et al. 1998, Heym et al. 1999). Molecular studies have demonstrated that mutations in katG and inhA (regulatory and structural regions) confer resistance to INH in Mycobacterium tuberculosis (Telenti 1998). Mutations in oxyR-ahpC intergenic region are considered a compensatory mechanism for the loss of KatG enzyme activity in resistant strains (Sherman et al. 1996, Ra-maswamy & Musser 1998) and can be used as a surrogate marker for detection of resistance to INH in M. tuberculosis (Wilson et al. 1998). Other putative mechanisms that can result in resistance to INH are less well understood (Zang & Telenti 2000, Larsen et al. 2002). Prior studies (Chen & Bishai 1998, Miesel et al. 1998) have postulated the involvement of the ndh gene in resistance to INH and ethionamide (ETH) in M. smegmatis. Mutations in this gene result in diminishment of NADH oxidation and consequently an increase in NADH concentration and depletion of NAD+. This alteration of the NADH/NAD+ ratio than prevents the peroxidation reactions required for the activation of INH, as well as the displacement of the NADH-isonicotinic acyl complex from InhA enzyme binding site. In order to evaluate the contribution of the ndh gene to INH resistance in M. tuberculosis, Lee et al. (2001) sequenced the entire ndh gene of 84 INH-resistant and 22 susceptible M. tuberculosis isolates. This study identified two distinct mutations in eight INH- resistant isolates (T110A and R268H), while no mutations were detected in susceptible isolates. Recently, ndh mutants of M. bovis BCG were shown to be low-level resistant to INH and ETH (Vilchèze et al. 2005). These mutants had increased intracellular NADH/NAD+ ratios and, upon complementation with the wild-type ndh gene, full susceptibility to both INH and ETH was restored. In this sense, we investigated the presence of ndh mutations among INH-resistant and susceptible M. tuberculosis clinical isolates from Brazil. We studied 23 INH-resistant and 13 susceptible M. tuberculosis clinical isolates from the culture collections of the Institute Adolfo Lutz of Ribeirão Preto, Institute Adolfo Lutz of Sorocaba, Institute Clemente Ferreira, state of São Paulo, and Clinical Bacteriology Laboratory, Department of Clinical Analysis, state of Paraná, Brazil. All isolates had previously been identified by biochemical test (Kent & Kubica 1985) and genotyped using the spoligotyping method (Molluizen et al. 1998). Susceptibility to INH was determined using the proportion method in Löwenstein- Jensen medium (BD/Becton, Dickinson and Co. Sparks, MD, US) according to Canetti et al. (1963). The minimal inhibitory concentration (MIC) of INH was determined for all isolates using the Microplate Alamar Blue Assay (MABA) (Franzblau et al. 1998). The isolates were cultured in Middlebrook 7H9 broth with oleic acid-albumin-dextrose-catalase (OADC) Enrichment (BBL/Becton, Dickinson) containing two-fold serial dilution of INH, ranging from 32.0 to 0.25 µg/ml. The visual MIC value was defined as the lowest INH concentration that prevented a color change of the Alamar Blue reagent (Accumed International, Westlake, OH, US) from blue to pink. A blue color in the well was interpreted as absent (inhibit grow-up of 90%) of mycobacterial growing. According to Franzblau et al. (1998), in this study, the M. tuberculosis isolates were considered susceptible to INH when showed MIC value ≤ 0.25 µg/ml. In all isolates studied here, an earlier report describes the genotypic analysis by polymerase chain reaction-single-strand conformation polymorphism analysis (PCR-SSCP) targeting the codons 16 (region - 7 to 67) and 94 (region 65 to 125) of inhA, the oxyR-ahpC intergenic region and the entire kasA gene by amplifying six overlapping fragments. Sequencing was performed of the genes fragments that presented PCR-SSCP patterns different from wild type ones and of the entire katG gene of all strains studied in this work (Cardoso et al. 2004). The chromosomal DNA was extracted as described earlier (Gonzalez-y-Merchand et al. 1996) with modifications introduced in our laboratory (Cardoso et al. 2004). The entire ndh gene was PCR amplified in five overlapping fragments according to Lee et al. (2001). The PCR products were generated using the Hotstart TaqTM Master Kit (Qiagen Inc. Valencia, CA, US) according to the manufacturer's instructions with 1 µl of template genomic DNA in a final reaction volume of 25 µl in a Gene Amp System 2400 thermalcycler (PE Applied Biosystems Corp., Foster City, CA, US). Sequencing reactions were performed using the ABI Prism BigDye terminator Cycle Sequencing Kit (PE Applied Biosys-tems Corp.) according to the manufacturer's instructions in a Gene Amp PCR System 9700 ther-malcyrcler (PE Applied Biosystems Corp.). Sequencing products were purified using CENTRI-SEP Colums (Princeton Separations, Inc. Adelphia, NJ, US) and electrophoresed on an ABI Prism 373XL automatic sequencer (PE Applied Biosystems Corp.). Sequence data were assembled and edited electronically using ABI PRISM DNA Sequencing Analysis Software version 3.0 (PE Applied Biosystem Corp.) and the results compared to the sequence for ndh of M. tuberculosis H37Rv present in GeneBank (acession no. Z838559). From the 23 INH-resistant M. tuberculosis clinical isolates studied, ndh gene mutations were detected in two (8.7%) and no mutation was detected in the 13 INH-susceptible isolates (Table). One isolate (INH MIC > 32.0 µg/ml) presented mutation in codon 13 (CGT to TGT), resulting in an R13C amino acid change and in other (INH MIC 32.0 mg/ml) the mutation was in codon 18 (GTG to GCG) resulting in a V18A amino acid change. The R13C mutations detected in our work occurred at different locations from those previously reported ndh either in M. smegmatis (Chen & Bishai 1998, Miesel et al. 1998) or in clinical isolates of M. tuberculosis (Lee et al. 2001, Ramaswamy et al. 2003) but the V18A mutation was reported by Ramaswamy et al. (2003) in susceptible M. tuberculosis isolates. It was verified that both INH-resistant isolates, that presented mutation in ndh, also had a mutation in katG (one of them with point mutation in codon no. 315, resulting in a S315T and in the other isolate, insertion of A at nucleotide position 17 of katG) detected in our previous study (Cardoso et al. 2004). According to Ramaswamy and Musser (1998), Zhang and Telenti (2000), van Soolingen et al. (2002), isolates with katG mutation, show MIC value consistent with of the INH-resistant isolates, suggesting that resistance was due to mutation in katG. In this sense, the contribution of ndh mutation to the INH resistance phenotype is unknown. Considering the recent demonstration that mutations in the ndh gene of M. bovis BCG mediate INH and ETH resistance (Vilchèze et al. 2005) it seems probable that such mutation could have a similar effect in M. tuberculosis. The R13C ndh mutants identified in this study might prove useful in demonstrating the involvement of this mechanism in M. tuberculosis. Further investigation is needed to establish the role and prevalence of ndh mutations in a larger number of INH-resistant M. tuberculosis clinical isolates. Two INH-resistant isolates studied here (INH MIC 2.0 and > 32.0 µg/ml) that showed no mutation in ndh genes also were negative to katG, inhA, kasA, or in the oxyR-ahpC intergenic region mutation in the previous study (Cardoso et al. 2004). This suggests the existence of additional INH resistance mechanism. These isolates are therefore good candidates for further investigation of alternative mechanisms. ACKNOWLEDGMENTS To Glenn Paul Morlock and Robert Cannon Cooksey (Division of AIDS, STD, and TB Laboratory Research, National Center for HIV, STD, and TB Prevention, Centers for Disease Control and Prevention, Atlanta, Georgia) for their collaboration in interpretation of data, suggestion, and critical comments on the manuscript. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07009t1.jpg] |
| |||||||||

{kind=link}