 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No.1, February 2007, pp. 83-85 Aspartic protease activities of schistosomes cleave mammalian hemoglobins in a host-specific manner Jeffrey W Koehler/*/**, Maria E Morales/**, Bryan D Shelby/*/**, Paul J Brindley/**/+ Department of Tropical
Medicine*, Microbiology & Immunology**, Interdisciplinary Program in
Molecular and Cellular Biology, Tulane University Health Sciences Center,
1430 Tulane Avenue, New Orleans, LA 70112, US Received 6 October
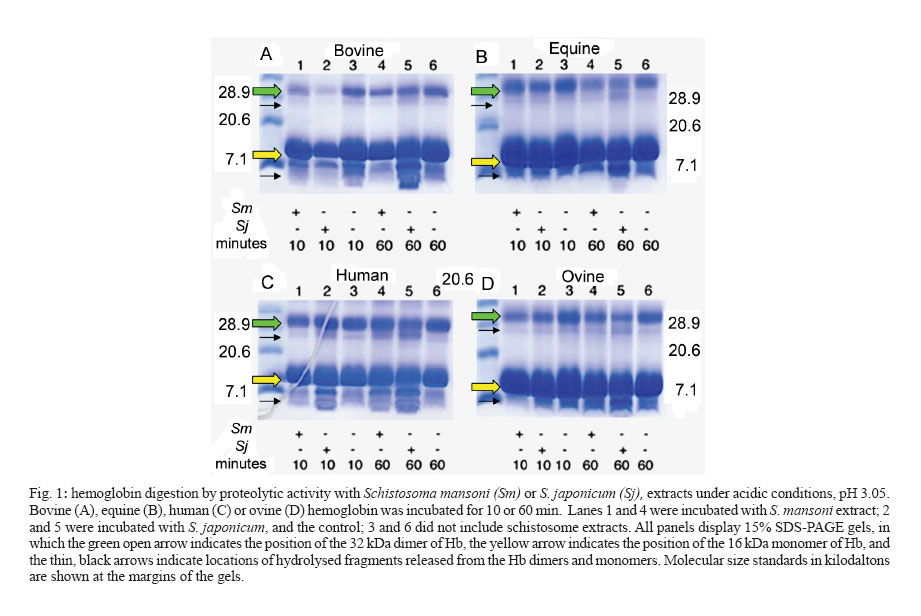
2006 Code Number: oc07014 We examined the efficiency of digestion of hemoglobin from four mammalian species, human, cow, sheep, and horse by acidic extracts of mixed sex adults of Schistosoma japonicum and S. mansoni. Activity ascribable to aspartic protease(s) from S. japonicum and S. mansoni cleaved human hemoglobin. In addition, aspartic protease activities from S. japonicum cleaved hemoglobin from bovine, sheep, and horse blood more efficiently than did the activity from extracts of S. mansoni. These findings support the hypothesis that substrate specificity of hemoglobin-degrading proteases employed by blood feeding helminth parasites influences parasite host species range; differences in amino acid sequences in key sites of the parasite proteases interact less or more efficiently with the hemoglobins of permissive or non-permissive hosts. Key words: schistosome - hemoglobin - aspartic protease - cathepsin D - host specificity - host range Every species of infectious organism exhibits a varying degree of host-specificity, a nebulous characteristic that essentially defines the species range of its potential hosts. Once a parasite makes intimate contact with its prospective host, physiological compatibility becomes the most critical, but still a poorly understood, determinant of the success of parasitism and thus of host-specificity. Among factors within the host that define physiological compatibility are the availability of appropriate and sufficient nutrients and suitable physical, chemical, and immunological conditions that allow the parasite to establish, mature, and reproduce. Moreover, once a host-parasite relationship has evolved, the parasite and the host are locked into an evolutionary arms race [e.g. The Red Queen Hypothesis (Lively & Dybdahl 2000)]. This is presumed to manifest at the molecular level in the form of finely tuned enzyme-substrate, receptor-ligand, and like molecular interactions. We hypothesize that the evolution of fine specificity of these molecular interactions at various stages of the infectious cycle influences the evolution of host range of a given parasite. Indeed, recent findings from our laboratories have shown that this hypothesis is supported, at least for several hemoglobin-degrading, aspartic proteases, using molecular modelling approaches (Brinkworth et al. 2000, 2001) and biochemical approaches in vitro with hookworm enzymes (Williamson et al. 2002, 2003a, b, c). To further explore this phenomenon, in this short communication we present findings of an investigation of the capacity of acid proteases of two species of schistosomes, Schistosoma japonicum and S. mansoni, to digest hemoglobin from four different mammalian species, which included both permissive and non-permissive host species of the two schistosomes. MATERIALS AND METHODS Schistosome extracts - Adult S. japonicum (from an Anhui, China isolate) and S. mansoni (NMRI strain from Puerto Rico) were obtained from experimentally infected mice, as described (Brindley et al. 2001). Soluble protein extracts from about 200 mixed sex adults of S. japonicum and S. mansoni were prepared by sonication-induced lysis (5 x 5 second bursts on ice, output control value 3, model W-220F Sonicator, Heat Systems, Ultrasonics Inc., Plainview, New York) of the schistosomes in 100 mM sodium phosphate, pH 7.0. After centrifugation of the lysate for 30 min at 4ºC at 14,000 rpm, the supernatant was employed as soluble schistosome extract. Protein concentrations of extracts and hemoglobin were determined spectrophotometrically using the BCA kit (Pierce, Rockford, IL). The concentrations of the extracts were 10.8 mg/ml and 9.1 mg/ml for the soluble extracts from S. japonicum and S. mansoni, respectively. Identical amounts of schistosome protein, 2.16 mg, were employed in each digestion reaction, as described below. Hemoglobins - Whole bloods from horse, sheep, and cattle were purchased from commercial sources. Human blood was donated by a healthy laboratory colleague. Hemoglobin (Hb) was prepared by lysing erythrocytes (which had been washed four times in 1 x PBS, pH 7.2 to remove the plasma) in hypotonic medium, 0.1 x PBS. The cell debris was removed by centrifugation at 14,000 x g for 30 min, and the supernatant was employed as Hb (Becker et al. 1995). Hb concentrations, determined as above, of the four mammalian species were 9.8 (human), 12.9 (ovine), 26.6 (equine), and 27.5 μg/μl (bovine), respectively. Proteolysis of hemoglobin - Hb (approx. 30 - 50 μg) was incubated with schistosome extract (2.16 mg protein) in sodium formate (pH 3.05) at 37°C for 10 min or 60 min. Aliquots of the resulting hydrolysates were mixed with nonreducing SDS-polyacrylamide gel electrophoresis sample buffer and separated by electrophoresis through 10 or 15% SDS-polyacrylamide gels, after which the gels were stained with Coomassie Brilliant Blue R-250. Protein standards (broad-range, Bio-Rad) were included in each gel. Images of the Coomassie-stained gels were documented using the Versa-doc system driven by Quantity 1 software (Bio-Rad). RESULTS AND DISCUSSION The three major species of human schistosomes exhibit differences in their preferred mammalian hosts. S. japonicum displays a broad host range in respect to the numbers of mammalian species in which it naturally infects and reproduces. S. mansoni and S. haematobium display much more limited host species ranges (see Brandt & Loker 2005). Mammalian stages of these human schistosomes reside within the blood vessels of their hosts, where they ingest blood as the major source of amino acid nutrients (Brindley et al. 2001, Caffrey et al. 2004, Delcroix et al. 2006). Here we examined the digestion of hemoglobin from four mammalian species, human, cattle, sheep and horse at highly acid pH by extracts of mixed sex adults of S. japonicum and S. mansoni. Extracts of the S. japonicum worms more efficiently digested the hemoglobin from bovine, ovine, and equine blood compared to the S. mansoni extract (Fig. 1A, B, D, lanes 4 compared to lanes 5 for the S. mansoni versus the S. japonicum extracts). Both species' extracts digested human Hb well (Fig. 1C). The trend, enhanced digestion of bovine, ovine, and equine Hb by S. japo-nicum compared to S. mansoni extracts, in general was apparent whether the digests were examined after 10 min or 60 min incubation (Fig. 1). More specifically, in the gels presented in Fig. 1, the Coomassie-stained monomer of Hb of ~ 16 kDa and the dimer of ~ 32 kDa are readily apparent; the position of the 16 kDa monomer is indicated with an open (yellow) arrow, and the position of the 32 kDa dimer is indicated with an open (green) arrow. (The monomer of 16 kDa is likely to be a mixture of fragments of the alpha and beta chains of Hb; the dimer is an alpha-beta heterodimer). After addition of schistosome extracts, degradation of the Hb monomer and dimer was indicated by an increase in lower molecular size products migrating faster than the 32 kDa dimers and 16 kDa monomers of Hb (on Fig. 1, thin black arrows indicate locations of these lower molecular mass degradation fragments). Further analyses undertaken with bovine Hb, using 10 or 15% SDS-PAGE gels produced similar findings to those presented in Fig. 1, in that the S. japonicum extract more thoroughly digested bovine Hb than did the S. mansoni extract (not shown). Because these digestions were carried out in highly acidic conditions, pH 3.05, the protease activity responsible for the digestion of the Hb can be ascribed to the gut-localized cathepsin D aspartic protease of S. mansoni and S. japonicum. This orthologue plays a key role in proteolysis of hemoglobin released from ingested erythrocytes (Brindley et al. 2001), indeed may make the earliest cleavages (Delcroix et al. 2006), and is the only aspartic protease known from schistosomes (Hu et al. 2003, Morales et al. 2004). Whereas there are other protease activities in the extracts of schistosomes, including additional gut-associated, hemoglobin degrading enzymes (cathepsins B1, L1, L2, and C, legumain, and leucine aminopeptidase), none of these other proteolytic enzymes is active at pH 3 (see Caffrey et al. 2004, McCarthy et al. 2004, Delcroix et al. 2006). One interpretation of the results presented here is that the S. japonicum protease has broader substrate specificity than that of S. mansoni, which parallels the broader host range of S. japonicum. S. japonicum naturally infects bovines, sheep, and horses, in addition to humans, whereas of these mammals only the human is a permissive host for S. mansoni. These findings indicate that a similar situation may exist in schistosomes to that in hookworms, where it has been demonstrated that the species-specific specificity of proteases could contribute to host range. In particular, the aspartic protease Na-APR-1 from the human hookworm Necator americanus more proficiently hydrolyses human Hb than canine Hb, and where the aspartic protease Ac-APR-1 from the dog hookworm Ancylostoma caninum more proficiently hydrolyses canine Hb than human Hb. (N. americanus does not infect dogs, and A. caninum does not mature to patency in humans.) Whereas it will be important to elaborate on the studies described here using active recombinant forms of the S. japonicum and S. mansoni cathepsin D aspartic proteases, these findings support the hypothesis of Prociv and co-authors (Brinkworth et al. 2000) and the findings of Loukas and co-workers (Williamson et al. 2002, 2003a, 2004) that substrate specificity of hemoglobin-degrading proteases employed by blood feeding helminth parasites influences parasite host species range. Differences in amino acid sequences in key sites of the parasite proteases likely interact less or more efficiently with the hemoglobins of permissive or non-permissive hosts. The S. japonicum and S. mansoni cathepsin D orthologues exhibit ~ 88% amino acid sequence identity in the mature region, and display several amino acid differences in active site residues and cleavage sites in human haemoglobin (Brindley et al. 2001, Brinkworth et al. 2001). ACKNOWLEDGEMENTS To Mary Duke of the Queensland Institute of Medical Research, Brisbane, Australia for Schistosoma japonicum worms. S. mansoni-infected mice were supplied by Dr Fred A Lewis through NIH contract NO155270. PJB is a recipient of a Burroughs Wellcome Fund scholar award in Molecular Parasitology. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07014f1.jpg] |
| |||||||||

{kind=link}