 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No.1, February 2007, pp. 111-115 Impairment of cell division of Trypanosoma cruzi epimastigotes Michele A Zacks Department of Pathology,
University of Texas Medical Branch, 301 University Boulevard, Galveston,
TX 77555-0609, US Financial support: American Heart Association, John Sealy Memorial Endowment Fund for Biomedical Research, American Health Assistance Foundation, the National Institutes of Health (AI053098-01, AI054578-01), the James W. McLaughlin Predoctoral Fellowship Fund, the Department of Pathology and the Graduate School of Biomedical Sciences at the University of Texas Medical Branch Received 18 May
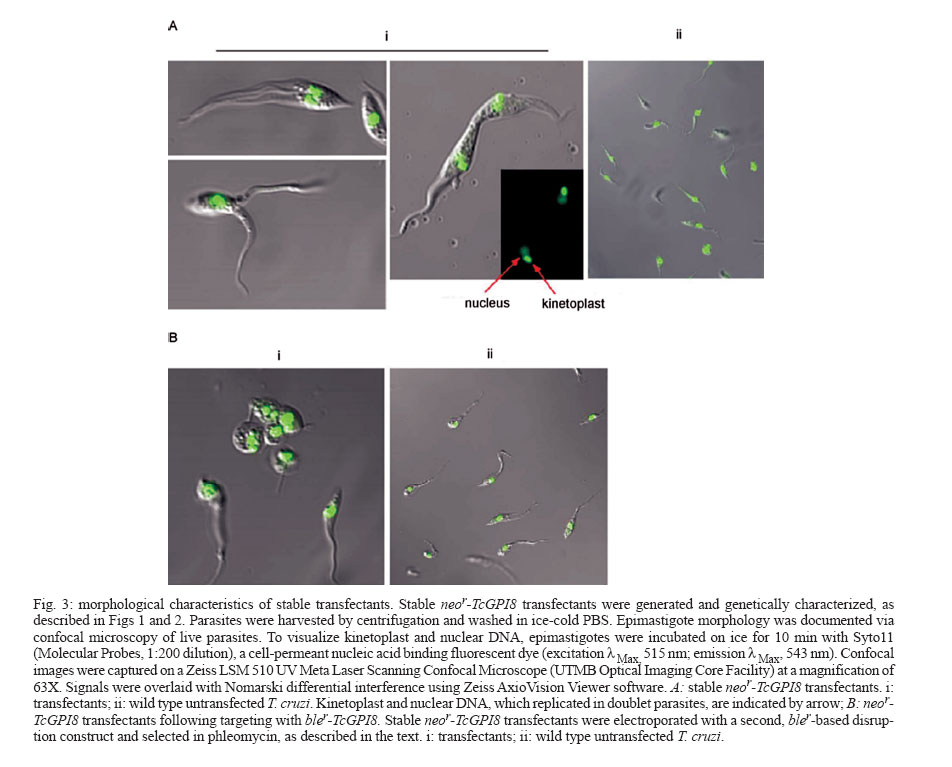
2006 Code Number: oc07019 The mechanisms that facilitate the adaptation of Trypanosoma cruzi to two distinct hosts, insect and vertebrate, are poorly understood, in part due to the limited ability to perform gene disruption studies by homologous recombination. This report describes a developmentally-defective phenotype that resulted from integration of a drug marker adjacent to the GAPDH gene in T. cruzi. Key words: Trypanosoma cruzi - life cycle - differentiation Trypanosoma cruzi is a protozoan parasite of the ancient branch of eukaryotes (Kingdom Eukaryota, Order Kinetoplastida) (Stevens et al. 1999) and is endemic in South and Central America, and Mexico. T. cruzi is transmitted to vertebrates, including humans, predominantly by insect vector (subfamily Triatoma, family Reduviidae) and also occurs via blood transfusion, organ transplantation, and congenital routes. In humans, 30% of chronically infected individuals are estimated to develop Chagas disease, a distinct form of cardiomyopathy (Miles 2003). Despite the description of the association between T. cruzi transmission via the triatomine insect vector and heart disease in 1909 and decades of research illuminating the extraordinary mechanism of host cell infection (Hall 1993, Burleigh & Andrews 1998), no vaccine is available and only two anti-parasitic drugs have been licensed for treatment. However, these drugs are effective mainly at the acute stage of infection and are highly toxic (Barrett et al. 2003). The mechanisms that facilitate the adaptation of T. cruzi to two distinct hosts, insect and vertebrate, are poorly understood, in part due to the limited ability to perform gene disruption studies by homologous recombination. This report describes a developmentally-defective phenotype that resulted from integration of a drug marker adjacent to the GAPDH gene in T. cruzi. This study was initiated to evaluate the role of surface expressed glycosylphophatidylinositol (GPI)-anchored proteins in the complex life cycle of T. cruzi and utilized a homologous recombination-mediated approach to targeted disruption of the TcGPI8 gene in the parasite, described in Zacks and Garg (2006) (Fig. 1). Electroporation with a neomycin-resistance (neor)-based construct targeting TcGPI8 and subsequent drug selection of transfectants resulted in genomic integration of the neor cassette, as confirmed by Southern blot analysis (Fig. 1) and PCR amplification of the 1.3 kilobase GAPDH-IR-neor fragment from gDNA of the trans-fectants (data not shown). However, GAPDHIR-neor did not integrate into the TcGPI8 gene and the 3' and 5' ends of TcGPI8 were absent (data not shown). Genome walking and PCR-cloning identified the site of insertion of the GAPDHIR/neor cassette (Fig. 2A-B) as adjacent to the 3' end of GAPDH (Fig. 2C) (Kendall et al. 1990). The orientation of the insertion is shown in Fig. 2D. The morphology of epimastigote-stage transfectants was documented via confocal microscopy (Fig. 3). Stable neor-TcGPI8 transfectants (Fig. 3A) showed unusual morphologies, first with the appearance of thin extended forms that appeared to be reduced in motility. Highly motile parasites with two or three flagella were present with frequent observation (> 2 per microscopy field) of two parasites with fused membranes, either in apical or longitudinal style. The nucleic acid staining pattern observed in doublet parasites incubated with the fluorescent Syto11 dye indicated that duplication of kinetoplast and nuclei was not impaired. However, in many cases, cellular membranes appeared to be fused. The possibility that these represent intermediate morphological forms in normal T. cruzi division cannot be excluded. However, such parasites were not found in either routine (e.g. daily) light microscope examination of non-transformed cultures during consideration over an extensive period of cultivation with selective drug (e.g., G418 for minimum of 2-3 months) or without selective drug while maintained in parallel cultures using liver infusion tryptose medium. Nor, as presented here, were such forms documented during confocal microscopy. This observation is consistent for this study as well as for cultures of pTEX transformed cultures that were continuously cultured for > 2 years in the presence of selective drug (e.g., G418, as reported here), under daily to weekly monitoring. At the time this phenotype became pronounced, growth of the transformant population declined and could not be further maintained, most likely as a result of their inability to complete their replication cycle. Thus, at the epimastigote stage, a striking defect in cell division occurred in transfectants. The effect of this transformation on the life cycle development of transfectants, e.g., differentiation of epimastigote to the infective metacyclic forms and infection of mammalian cells, was further evaluated. Transfectant epimastigotes and, as control, wild type epimastigotes were grown in liver infusion tryptose medium for ≥ 10 days without addition of media to achieve stationary-growth phase for the conversion of parasites to metacyclic trypomastigotes and were used to infect fibroblast monolayers at a parasite to cell ratio of 50:1. At 24 h after infection, medium containing free parasites was replaced and cells monitored for the appearance of trypomastigotes. Three independent in vitro infection experiments were performed in two different fibroblast cell lines (C2C12 and BHK21) using stationary-phase cultures of stable transfectants or of wild type untransfected T. cruzi. Transfectants did not convert to the typical morphology of the infective metacyclic form following ³ 10 days of cultivation without addition of new growth medium. No trypomastigotes were seen when monitoring fibroblasts infected with the stationary phase cultures of these neor-TcGPI8 transfectants during a 10-12 day incubation period. However, wild type, untransfected parasites were capable of infecting either fibroblast line, with appearance of trypomastigotes and amastigotes within 6 and 11 days, respectively. In summary, morphological transformation of the neor-TcGPI8 transfectant epimastigotes into metacyclic trypo-mastigotes in culture conditions of "starvation" did not occur, unlike for wild type parasites. In conclusion, these stable neor-TcGPI8 transfectants were not infective for mammalian cells in vitro. In effort to obtain homozygous transfectants, the stable neor-TcGPI8 transfectants were subsequently electroporated with a similar disruption construct containing the phleomycin-resistance (bler) gene in place of the neor gene. During drug selection (500 µg/ml ble), drastic changes from normal epimastigote morphology in axenic cultures were observed in these transfectants (Fig. 3B, panel i) but not in wild type parasites, which were cultured in parallel without selective drug (Fig. 3B, panel ii). Amastigote-like forms with short, retracted flagella were abundant (> 50% of the culture) among the transfected epimastigote population (Fig. 3B, panel i), whereas the morphology of the parallel wild-type untransfected culture was normal (Fig. 3B, panel ii). Uptake of propidium iodide in these forms indicated that parasite membranes were disrupted (data not shown). These parasites could not be maintained in culture, precluding isolation of gDNA for further genetic analysis. In summary, this report documents an interesting and selective defect in which organelle duplication (kinetoplast and nuclei) does not appear to be blocked, in which the flagella replicate but in some cases, in which the plasma membranes do not appear to separate effectively. From these studies, it can be surmised that cell division of epimastigotes was impaired selectively. To date, few studies have been published on the mechanism of cell division of T. cruzi (Gomez et al. 1998, 2001, Bogitsh et al. 1999, Grellier et al. 1999a, b, Orr et al. 2000, Santori et al. 2002). The pattern of development, e.g., the time course of DNA replication, organelle duplication and cell division of T. cruzi cannot be assumed to occur as has been demonstrated in elegant studies of the related trypanosomatid, T. brucei (Hendriks et al. 2000, McKean 2003). In T. cruzi, targeted disruption of genes for functional analysis remains a challenge and inducible expression systems have not been possible (DaRocha et al. 2004). In addition to the deficit of tools for genetic manipulation of T. cruzi, we lack an understanding of the mechanisms regulating T. cruzi gene expression and governing the cellular processes that enable the parasite to transform morphologically. However, recently, the genome sequence of T. cruzi CL-Brener strain was completed (El-Sayed et al. 2005). It is anticipated that future developments in transformation, gene disruption and/or inducible expression systems, combined with a comparative genomics approach (Parsons et al. 2005) will enhance our understanding of the complex developmental cycle and morphological transformation required for T. cruzi survival and may identify specific drug targets. Further molecular-genetic studies will be required to evaluate the potential role of the flanking sequences of GAPDH, such as promoter functions, in the regulation of cell division and differentiation in T. cruzi. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07019f3.jpg] [oc07019f2.jpg] [oc07019f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}