 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No.2, March 2007, pp. 191-195 Biomphalaria tenagophila potencial vector of Schistosoma mansoni in the Paraná River basin (Argentina and Paraguay) C Edgardo Borda+, María Josefa F Rea Centro Nacional
de Parasitología y Enfermedades Tropicales, Facultad de Medicina,
Universidad Nacional del Nordeste, 3400 Corrientes, Argentina Financial support: Secretaría General de Ciencia y Técnica, Universidad Nacional del Nordeste, Corrientes, Argentina Received 23 October
2006 Code Number: oc07033 Susceptibility and compatibility experiments were carried
out with 700 Biomphalaria tenagophila from the Paraná River
basin exposed to infection with Schistosoma mansoni. Individual infection
was performed with 10 miracidia of SJ2 strain from the Paraiba
valley (Brazil) originally infective to B. tenagophila. These snails
were laboratory-breed progeny of B. tenagophila collected from six
localities of Argentina and one from Paraguay. From Argentina: Rincón
de Vences (7%) and Posadas (11%) became infected with S. mansoni and
the calculation of Frandsen's index (TCP/100) shows that they were Class
II poorly compatible. Those snails from Goya (22%), Maloyas (5%), and Berón
de Astrada (3%) were Class III compatible to the S. mansoni. None of
the 100 snails exposed from Caá-Catí became infected (Class
0 incompatible). Tested samples from Paraguay (Encarnación) were infected
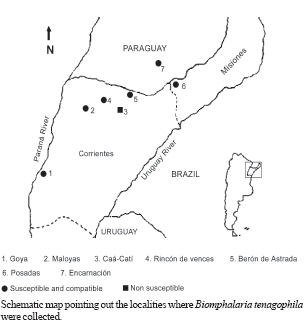
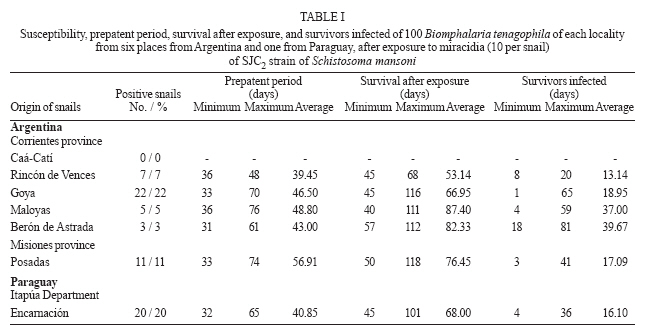
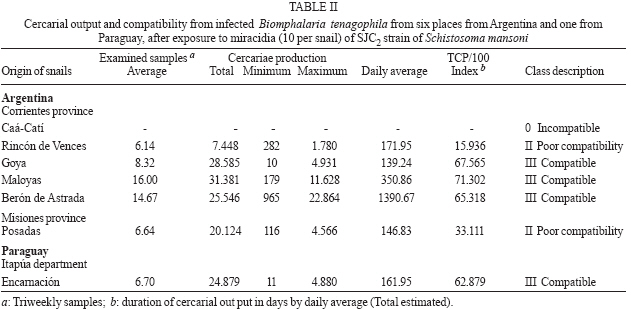
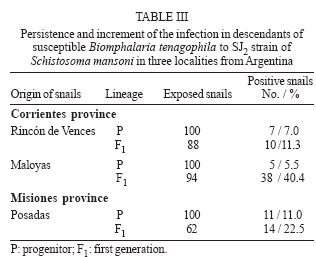
(20%) and compatible (Class III). In some America countries where the schistosomiasis is endemic the geographic distribution of the genus Biomphalaria, is very important for the prevention in vulnerable areas those present risk of transmission (Teles et al. 1989, 1991, Carvalho et al. 1997, 1998). Three mollusc species are recognized as Schistosoma mansoni intermediate hosts in Brazil: B. glabrata, B. tenagophila, and B. straminea (Paraense 1986). Argentina and Paraguay are in the Paraná River basin and they are areas still free from schistosomiasis. For that reason the knowledge of the planorbid snails and the species that could be considered potential intermediate hosts of S. mansoni is important for an appropriate epidemiological surveillance (Silva et al. 1994). B. tenagophila and B. straminea exist in the Paraná and Uruguay River basins in Argentina: Misiones, Chaco, Corrientes, and Entre Ríos; in Paraguay: Itapúa and Las Misiones; in Uruguay: Artigas and Salto (Borda & Rea 1995). B. aff. straminea from Espinillar near Salto, Uruguay, shows a significant level of susceptibility to infection with the SJ2 strain of S. mansoni (Paraense & Correa 1989). In previous searches Borda and Rea (1997) demonstrated that B. tenagophila from Ayolas (Paraguay) and from Fuente Salto (Uruguay) exposed to the SJ2 strain of S. mansoni were infected. The possibility of the expansion of schistosomiasis to Argentina and Paraguay where B. tenagophila occurs must be contemplated. However, investigation about the susceptibility of allopatric populations of B. tenagophila from the Paraná River basin to S. mansoni from the Paraiba valley (Brazil), the degree of compatibility between parasite and intermediate host and the persistence of the infection in their descendants is now reported. MATERIALS AND METHODS Malacological surveys were undertaken in six localities from Argentina Northeast and one from Paraguay Southeast (27º50' N and 52º50' W, Fig. 1). In Argentina the snails were collected in the Corrientes province: Caá-Catí, Rincón de Vences, Goya, Maloyas, and Berón de Astrada and from Posadas (Misiones province). The specimens from Paraguay proceeding from Encarnación (Itapúa Department) The snails of each locality were placed into an aquarium for rearing which were kept at a room temperature of 24-27°C. The descendants were considered of the progenitor generation and ten of these snails were identified with the techniques detailed in a previous work (Borda & Rea 1997). These descendants were infected with S. mansoni of SJ2 strain, from São José dos Campos, São Paulo, Brazil, more adapted to B. tenagophila (Paraense & Corrêa 1989). This strain is routinely maintained at the Cenpetrop laboratories by passages through sympatric B. tenagophila and hamster (Mesocricetus auratus) or albino mice (Mus musculus). One hundred specimens of each locality were individually exposed to 10 miracidia for at least 8 h. Snails with 4-8 mm diameter in size were used in all experiments, at the moment of infection. The procedures for exposure of snails to miracidia were those described by Paraense and Corrêa (1989). After 28 days, the molluscs from each group were individually exposed to artificial light 28-30°C (Standen 1952) and examined under stereomicroscope, to detect snails which were shedding cercariae. They were then examined weekly until 90 days after exposure, when negative snails were dissected and examined for development stages of the schistosome. The emission of cercariae was followed until death in all the positive specimens. Each infected snail, kept separately in an aquarium, was exposed to light, three days in the week, from 9 AM to 5 PM and the cercariae were counted (Paraense & Correa 1989). The total count for each snail was divided by the number of examined samples to find the average number of cercariae per sample. The product of this individual average by the duration in days of cercariae output gave an estimate of the total number of cercariae shed by that snail if daily counts had been made (Frandsen 1979 a,b). For comparison of infection rates, 100 B. tenagophila sympatric, were exposed each to 10 miracidia and followed up as described above, except for counts of cercariae. The F1 of infected snail from three localities from Argentina was obtained: Rincón de Vences, Maloyas, and Posadas. For the evaluation of the persistence of the susceptibility 62 to 94 snails F1 were used. For this purpose each snail was individually exposed to 10 miracidia. The results obtained were statistically analyzed using the Pearson test. RESULTS Of the 700 B. tenagophila exposed to SJ2 miracidia, the infection rate varied widely according to the snail populations, while all specimens from Caá-Catí proved unsusceptible to infection, the snails from six localities, five from Argentina and one from Paraguay became infected (Table I). The infection rate was between 3 and 22%. The prepatent period ranged from 31 to 76 days. The earliest shedding of cercariae was observed in one sample from Berón de Astrada on the 31st day of post-exposure: 43 ± 15.87 (mean and standard deviation). In the samples of other localities the onset of shedding occurred: in Encarnación (49.85 ± 10.81); Goya (39.68 ± 10.52); Posadas (56.91± 17.51); in those from Rincon de Vences (39.45 ± 5.86) and Maloyas (48.8 ± 16.02) From B. tenagophila controls, 20% of the exposed was infected and the prepatent period varied from 29-86 days. Snails from Goya and Encarnación showed susceptibility rate to S. mansoni infection superior to 20%, similarly to that of B. tenagophila controls. The shortest period of survival of the snails after exposure was 40 days observed in one specimen from Maloyas and the maximum of 118 days in one snail from Posadas. Survival after onset of cercarial output varied between 1 to 81 days. One snail from Berón de Astrada shed cercariae for 81 days and showed the highest production, both in total output and daily average (Tables I, II). Table II shows some data on cercarial output and compatibility concerning the 68 snails that shed cercariae. The snails from Rincón de Vences showed an index of TCP/100 of 15,936 cercariae (Class II poorly compatible). Also with the same parameters (index of TCP/100 of 33,111) the snails from Posadas are included in the same class. On the contrary, the snails from Goya, Maloyas, Berón de Astrada, and Encarnación were included in the Class III (compatible) with an index TCP/100 of more than 50,000 cercariae. No snail cured spontaneously of the infection. However, in two snails from Goya and one of Berón de Astrada the emission of cercariae was interrupted during two and 12 days, 20 days before the snail death. In Table III is shown the persistence of the infection in the F1 generation of susceptible B. tenagophila. Of the 244 B. tenagophila of the first generation (F1) from Rincón de Vences, Maloyas, and Posadas exposed to S. mansoni, 62 snails were susceptible. When we compared the susceptibility of progenitors no significant differences (χ2 = 2.43; p = 0.29) could be observed in the snails from Rincón de Vences, Maloyas, and Posadas. The same occurred when the infection rates of progenitors and F1 from Rincón de Vences were analyzed (χ2 = 0.89; p = 0.35). In contrast, significant differences occurred between the snails from Maloyas (χ2 = 27.22; p = 0.0001) and Posadas (χ2 = 4.24; p = 0.04) showing the increase of the infection rates from the progenitor to the F1 generation. DISCUSSION Foci of active transmission of schistosomiasis have appeared in Brazil in areas till then free from the disease, with serious difficulties for the control (Coura & Amaral 2004). Between 1981 and 1991, the endemia reached the locality from São Francisco do Sul and Jaraguá do Sul in the southeast, in the state of Santa Catarina (Bernardini & Machado 1981, Espinola et al. 1992) and B. tenagophila was the natural intermediate host. The southern limit of the geographical distribution of B. glabrata in Brazil before 1986 was Curitiba (25°25'S) in the state of Paraná (Paraense 1986). In 1997 a population of B. glabrata was found in Esteio, a locality of Rio Grande do Sul near Porto Alegre (Car-valho et al. 1998). Graeff-Teixeira et al. (1999) found foci of S. mansoni transmission next to metropolitan area of Porto Alegre (29°49'37''S, 5l°10'56''W), the capital of the state of Rio Grande do Sul and actually it is distinguished as a low endemicity area for the schistosomiasis expansion (Graeff-Teixeira et al. 2004). The states of Santa Catarina and Rio Grande do Sul border the Argentina Northeast and they are in the influence area of Rio de la Plata basin. In previous susceptibility experiments, Borda and Rea (1997) exposed B. tenagophila of five localities to three S. mansoni strains: snails from Caá-Catí (SJ strain); Rincón de Vences, Goya, and Encarnación (BH2 strain adapted to B. glabrata) and Posadas (SJ2 strain). None of the snails was infected. In this experience, snails of the mentioned localities, with a higher number of samples, were infected by the SJ2 strain. Only the snails from Caá-Catí were negative to the same strain. The behavior of those snails to the possible infection with S. mansoni strain which infect naturally two different species of Biomphalaria is identical to the resistance observed with B. tenagophila from Puerto Hormiguero, on the Uruguay River in the province of Corrientes (Pellegrino et al. 1968, Borda & Pellegrino 1976, Paraense & Correa 1978). This is the second opportunity that snails of that species were found naturally refractory to the infection in the province of Corrientes. In Brazil, it has been demonstrated that B. tenagophila possesses a wide range of natural and experimental infection rates with that parasite. During many years it was not recognized as natural intermediary host of S. mansoni. In his early studies Lutz (1916) referred to the insusceptibility of Planorbis nigricans (a synonym of B. tenagophila) and for that reason, he proposed in 1923 the name of Planorbis inmunis. The role of B. tenagophila as natural vector was demonstrated in Brazil in the state of São Paulo (Paraense & Deslandes 1956). The susceptibility of B. tenagophila to infection with the SJ strain was demonstrated in 15 regions of Brazil (Paraense & Correa 1978). The infection rates varied between 0.8 and 91.5%. But in the same survey three snail populations from Uraí (Paraná), Ubá (Minas Gerais), and Cuiabá (Mato Grosso) were refractory. The highest experimental rate was achieved by Correa dos Reis et al. (1979) with 100% of infection with B. tenagophila from Cabo Frio (Rio de Janeiro). B. tenagophila Taim, from the Ecological Station of Taim, state of Rio Grande do Sul, Brazil, proved to be systematically resistant to S. mansoni. This resistance is a dominant character and could be succesful to obtain a model of control in areas where the transmission of schistosomiasis is only due to B. tenagophila (Coelho et al. 2004). The degree of compatibility between the parasite and the snail was determined for a better evaluation of the epidemiological importance of that specie in a region free of schistosomiasis. However, we developed a quantitative method TCP/100 index (degree of compatibility between the Schistosoma and its intermediary host) proposed by Frandsen (1979 a,b). Of six populations of B. tenagophila under investigation, the snails from Posadas and Rincón de Vences were poorly compatible to the SJ2 strain (Class II), index less than 50,000 cercariae. Meanwhile, other four populations from Goya, Maloyas, Berón de Astrada, and Encarnación were compatible with a higher index of 50,000 (Class III). This compatibility (Class III with SJ strain) is similar to that reported in B. tenagophila from Pampulha Lake in Belo Horizonte (Minas Gerais, Brazil) by Souza et al. (1995). Snails of that species naturally infected with S. mansoni were already found in several opportunities in the Pampulha Lake (Souza et al. 1987). It was demonstrated once again, that the transmission of the trematode depends of compatibility of S. mansoni strain (Locker & Bayne 1986), in this case of experimental character. Newton (1953) was the first in to prove the hereditary nature of the susceptibility from B. glabrata to infection with S. mansoni. Later on, other authors also suggested the participation of bound genetic factors to the infection of that planorbids species (Richards & Merrit 1972, Richards 1975). Santana et al. (1978) obtained populations of B. glabrata and B. tenagophila highly susceptible to the infection with the BH and SJ strain of S. mansoni after the selection for autogamy of susceptible snail descendants. The results of these experiences suggest the genome of some B. tenagophila populations, genetic variants to certain S. mansoni strain exists. When the miracidia penetrated in the snails, some resistance or compatible variant will be found and, as the susceptibility is an inheritance character, when the snail offspring is exposed to the same parasite strain, this will select the growth of the susceptible variant. These observations about the persistence of the infection were demonstrated in B. tenagophila from Rincón de Vences, Maloyas, and Posadas, with a similar procedure of autogamy selection of B. tenagophila susceptible, even the infection rate with SJ2 strain in the samples F1 from Maloyas and Posadas was increased in connection with the progenitor snails (Table III). Data on susceptibility presented in this paper strongly suggest the possibility of B. tenagophila coming to act as a vector of S. mansoni in the Argentina Northeast. The possibility of introduction and spread of schistosomiasis in the Rio de la Plata basin is a potential risk in consequence of the construction of roads and dams, migration and movement of people from endemic areas from Brazil with faulty sanitary conditions. ACKNOWLEDGEMENTS To Dr Wladimir Lobato Paraense, Instituto Oswaldo Cruz, Rio de Janeiro, Brazil for S. mansoni strain. To Cristina Gené, Osvaldo Benitez, and Luis Armando Mosqueda, staff of Schistosomiasis Laboratory of Cenpetrop. To Amalia Nélida C de Contreras (Conicet) for assistance with the English in this manuscript. To Dr Silvia Mazza, Facultad de Ciencias Agrarias, Universidad Nacional del Nordeste, Corrientes for statistical analysis. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07033t3.jpg] [oc07033t2.jpg] [oc07033t1.jpg] [oc07033f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}