 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No.2, March 2007, pp. 197-202 Detection of Onchocerca volvulus (Nematoda: Onchocercidae) infection in vectors from Amazonian Brazil following mass Mectizan™ distribution Verônica Marchon-Silva/+, Julien Charles Caër*, Rory James Post**, Marilza Maia-Herzog, Octavio Fernandes* Laboratório
de Referência Nacional em Simulídeos e Oncocercose, Departamento
de Entomologia *Laboratório de Epidemiologia Molecular de Doenças
Infecciosas, Departamento de Medicina Tropical, Instituto Oswaldo Cruz-Fiocruz,
Av. Brasil 4365, 21040-900 Rio de Janeiro, RJ, Brasil **Department
of Entomology, The Natural History Museum, London, U.K. Financial support: Fundação Nacional da Saúde/RO, CDTV and CGLab/SV/MS, Instituto Oswaldo Cruz, Onchocerciasis Elimination Program for the Americas Received 31 October
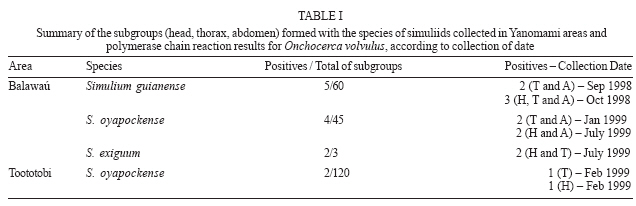
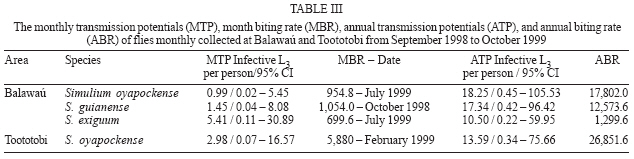
2006 Code Number: oc07034 Detection of Onchocerca volvulus in Simulium populations is of primary importance in the assessment of the effectiveness of onchocerciasis control programs. In Brazil, the main focus of onchocerciasis is in the Amazon region, in a Yanomami reserve. The main onchocerciasis control strategy in Brazil is the semi-annually mass distribution of the microfilaricide ivermectin. In accordance with the control strategy for the disease, polymerase chain reaction (PCR) was applied in pools of simuliids from the area to detect the helminth infection in the vectors, as recommended by the Onchocerciasis Elimination Program for the Americas and the World Health Organization. Systematic sampling was performed monthly from September 1998 to October 1999, and a total of 4942 blackflies were collected from two sites (2576 from Balawaú and 2366 from Toototobi). The molecular methodology was found to be highly sensitive and specific for the detection of infected and/or infective blackflies in pools of 50 blackflies. The results from the material collected under field conditions showed that after the sixth cycle of distribution of ivermectin, the prevalence of infected blackflies with O. volvulus had decreased from 8.6 to 0.3% in Balawaú and from 4 to 0.1% in Toototobi. Key words: Simulium - Onchocerca volvulus - polymerase chain reaction - onchocerciasis - ivermectin - Brazil According to the World Health Organization (WHO 2000) more than 120 million people, worldwide, live under risk of acquiring onchocerciasis and 18 million are infected. One third of those infected manifest dermatitis and 500,000 present serious visual problems with approximately 1% developing blindness. Indeed, 270,000 individuals are blind from this etiology worldwide. This disease is caused by infection with Onchocerca volvulus (Nematoda: Onchocercidae) and is endemic to Africa, the Arabian Peninsula, and Latin America, where it occurs in parts of Brazil, Colombia, Ecuador, Guatemala, Mexico, and Venezuela. In Brazil, the only known active focus of the disease is located in the Amazonian region, and extends across the border to form the southern focus of Venezuela. It corresponds to more or less the whole territory of the Yanomami in the northwest of the state of Roraima and central north of the state of Amazonas (Shelley 1991, 2002), exposing 13,767 indians to the infection (Funasa 2002). In the New World the control program has been sponsored by the Onchocerciasis Elimination Program for the Americas (OEPA). This program has as its main strategy the elimination of the disease through the semi-annually distribution of the ivermectin (Mectizan™) for 10 to 15 years to the population at risk (Blanks et al. 1998). This approach has been demonstrated to result in a decrease of microfilariae rates in the skin which gives immediate clinical benefit and reduces the possibility of infecting new vectors which is expected to reduce or eliminate transmission (Greene et al. 1989, Duke 1990). In simuliids, the classic parasitological method for the parasite detection (by dissection and examination of vectors) is unsuitable for high throughput analysis because it is time consuming and requires high levels of training, for example it requires the accurate morphological identification of the larvae. Since the 1990s, the polymerase chain reaction (PCR) amplification of an Onchocerca-specific repeated DNA sequence (O-150) has been used to detect and identify O. volvulus (Meredith et al. 1991, 1994, Zimmerman et al. 1992, 1993, Fischer et al. 1996, Merriweather & Unnasch 1996, Unnasch & Meredith 1996). However, in populations of vectors with a low prevalence of infection (as would be expected during an onchocerciasis control program using ivermectin, for example) large numbers of flies would have to be processed to obtain an accurate estimate of infection, which would be time-consuming and expensive. However, the O-150 PCR has proved to have a high analytical sensitivity, being able to detect a single infected fly in pools containing up to 100 flies (Katholi et al. 1995). Therefore, a statistical model has been developed by Katholi et al. (1995) to determine the prevalence of infection in the simuliid population, based on the prevalence of infection in pools of vectors, and it has been demonstrated that this method reveals a prevalence of infected flies statistically similar to the rates obtained by traditional entomological dissection and parasite microscopic examination (Yamèogo et al. 1999, Rodríguez-Pérez et al. 1999). In this study the aforementioned method was applied to evaluate the O. volvulus infection rate of pools containing 50 blackflies from Balawaú and Toototobi, two Yanomami tribal areas located in the Brazilian focus of onchocerciasis. MATERIALS AND METHODS Study sites and simuliids collection - Entomological collections were conducted in two distinct areas: (i) a hyperendemic area - Balawaú (01o48'39''N, 63o47'10"W; altitude of 340 m - northwest state of Amazonas) and (ii) a mesoendemic area - Toototobi (01o45'51"N, 63o37'28"W; altitude of 180 m - northwest state of Amazonas) (Figure). These sites are sentinel regions under surveillance of the Onchocerciasis Control Program of the Amazonian Focus/Brazil suggested by the National Foundation of Health (Funasa). The Mectizan™ distribution has been carried out annually in 1995 and 1996, and semi-annually in these areas since then. During the collection of the entomological specimens; the 6th, 7th and 8th cycles of treatment were implemented. Appetitive flies were collected at human bait over three consecutives days monthly from September 1998 to October 1999 at both Balawaú and Toototobi. The capture of the specimens started at 7:00 a.m. and ended at 7:00 p.m., consisting of 50 min sampling units followed by 10 min break. The host-seeking specimens were captured upon alighting and before beginning to feed on the collectors. To ensure that none of the captured flies had been able to ingest microfilariae from the collector, all flies showing evidence of recent blood meal were discarded (Davies et al. 1998). The flies collected were preserved in absolute ethanol (Post et al. 1993) until processed for DNA recovery in pools of 50 insects. Identification and preparation of pools - Taxonomic identification was carried out at the National Reference Laboratory for Simuliidae and Onchocerciasis, Oswaldo Cruz Foundation (Fiocruz) according to Shelley et al. (1997). Flies were divided in pools of 50 specimens according to the species, place, dates, and time of capture. To determine the prevalence of both infected and infective flies, all the specimens from each pool were divided by mechanical dissection in head (H), thorax (T), and abdomen (A) and these sub-sample pools stored separately. DNA extraction - The pools were placed in 1.5 ml snap-cap microcentrifuge tubes, and rinsed three times in 95% ethanol. Ethanol was removed using a pipette and the remainder allowed to evaporate for about 10 min at room temperature (RT). Separated heads, thoraxes, and abdomens were submitted to freezing in liquid nitrogen and macerated with pestles. Pools were homogenized in 300 μl of lysis buffer (100 mM NaCl; 10 mM Tris-HCl pH 8.0; 1 mM ethylenediamine tetra-acetic acid - EDTA; 0.1% sodium dodecyl sulphate - SDS) with 2 μl salmon sperm (250 ng/ml) and 100 μl of proteinase K (400 mg/ml). The mixtures were incubated at 55ºC for 1 h, and then boiled for 30 min with 4 μl of 1M dithiothreitol (DTT). The homogenates were submitted to two cycles of freezing in liquid nitrogen and thawing at RT and then another 1 μl of salmon sperm (250 ng/ml) was added. The mixture was extracted twice with phenol: chloroform:isoamilic alchool (25:24:1). A final extraction with chloroform was performed. After this procedure DNA was purified by adding three volumes of NaI solution (90.3 g sodium iodide; 1.5 g NASO3) and 5 μl of glass slurry (Sephaglas bond prep kit, Pharmacia Biotech®). The mixtures were agitated vigorously, placed at 4ºC for 15 min and centrifuged for 30 s. The glass slurry sediments were washed three times with 500 μl of ethanol wash solution [10 mM Tris-HCl pH 7.5; 100 mM NaCl; 1 mM EDTA, 50% (v/v) ethanol]. The pellets were dried at 35ºC for 20 min and resuspended in 100 μl TE (10 mM Tris-HCl pH 8.0; 1 mM EDTA pH 8.0). PCR amplification - Five μl of DNA from pools of heads, thoraxes, and abdomens were used in PCR reactions with 1X enzyme buffer (100 mM Tris-HCl, pH 8.3; 500 mM KCl, 6 mM MgCl2), 0.2 mM dNTPs (Perkin Elmer), 0.5 mM of each primer 1: 5'-GATTYTTCC-GRCGAAXARCGC-3' and 5'-GCXRTRTAAATXTG-YAAATTC-3' (Nutman et al. 1994, Unnasch & Meredith 1996), 1U of Taq polymerase (Applied Biosystems) and 2.5% formamide (Sarkar et al. 1990). The positive controls consisted of 1 ng of plasmid DNA pOVS134 (Meredith et al. 1989 - final concentration 20 pg/ml) and in the negative controls, 5 μl of distilled water were used instead of DNA. PCR conditions were one cycle of denaturation at 95oC for 10 min; 35 cycles of 1 min at 94oC, 2 min at 37oC, and 30 s at 72oC. A final step of 72oC for 5 min was performed. The PCR products were submitted to 2% agarose gel electrophoresis (Zimmerman et al. 1992) and visualized with ethidium bromide (10 mg/ml). Alternatively, 10% acry-lamide gel electrophoresis and silver staining was used (Kit DNA Silver Staining - Pharmacia). Insects used as negative controls - In order to have uninfected adult simuliids to be used as negative controls, pupae were collected and maintained in a humid container until adults emerged. These insects were collected from known breeding sites of Simulium oyapockense (Cachoeira do Bem Querer, Caracaraí, RR) and S. incrustatum (Igarapé Murupú, Boa Vista, RR), both outside the focus. Statistical analysis - The software Poolscreen (Katholi et al. 1995) was used to determine the overall prevalence of infected and/or infective flies using a 95% confidence interval (CI). The infection rate was determined by any positive PCR in any sub-group (head, thorax or abdomen), but the prevalence of infective flies was calculated only from the pools of heads. The monthly and annual transmission potential (MTP and ATP) is the measure of the number of infective larvae that a person who resides in an O. volvulus endemic area is exposed (L3 per person) and were also calculated with the same CI. RESULTS Simuliids collection and identification - The total number of flies collected was 4942. From Balawaú 2576 were collected and identified as S. oyapockense (n = 1459), S. guianense (n = 1021), and S. exiguum (n = 105). From Toototobi 2366 were collected and all of them were identified as S. oyapockense. Simuliids from Balawaú were clustered into 36 pools (108 subgroups of head, thorax, and abdomen) - 20 of S. oyapockense, 15 of S. guianense, and 1 of S. exiguum. From Toototobi, 40 pools of S. oyapockense were formed (120 subgroups). Diagnosis of infectivity/infection in simuliids pools from Balawaú and Toototobi - Eleven subgroups (11/108) formed by head, thorax, and abdomen from insects collected in Balawaú were positive - 5 of S. guianense, 4 of S. oyapockense, and 2 of S. exiguum. All the 11 positive subgroups presented consistent results. Regarding S. guianense, two were from the analysis of the thorax and abdomen of the insects that comprised one pool captured in September 1998. Three were positive for the head, thorax, and abdomen subgroups from another pool collected in October 1998. The analysis of S. oyapockense subgroups revealed that two of the positive subgroups (thorax and abdomens) were from the pool captured in January 1999. The other two subgroups correspond to heads and abdomens belonging to another pool captured in July 1999. The two positive subgroups of S. exiguum correspond to heads and thorax of insects of the same pool captured in July 1999 (Table I). Two subgroups (2/120) of segments of S. oyapockense collected in Toototobi were positive corresponding to the thorax of one pool and to the heads of another pool, both collected in February 1999 (Table I). Statistical analysis - In Balawaú, the prevalence of infection determined by the Poolscreen Program was 0.30% (95% CI: 0.00096 0.00697, n = 36 pools of 50 flies; with 31 negative pools), being 0.21% for S. oyapockense (n = 20; with 18 negative pools - 95% CI: 0.0003 0.00760), 0.29% for S. guianense (n = 15; with 13 negative pools - 95% CI: 0.00033 0.01031) (Table II). The overall prevalence of infective flies was 0.17% (n = 36 pools; with 33 negative pools - 95% CI: 0.00035 0.00508), being 0.10% for S. oyapockense (n = 20; with 19 negative pools - 95% CI: 0.00003 0.00570), 0.14% for S. guianense (n = 15; with 14 negative pools - 95% CI: 0.00003 0.00767) (Table II). The number of flies classified as S. exiguum was only sufficient for a single pool. When this pool was divided into the three subgroups, the subgroups composed of heads and thoraxes were both positive. The highest MTPs were: (i) for S. oyapockense 0.99 infective larvae per person (95% CI: 0.02 5.45 L3/person; month biting rate (MBR) - July 1999 = 954.8); (ii) for S. guianense 1.45 infective larvae per person (95% CI: 0.04 8.08 L3/person; MBR - October 1998 = 1,054.0) and for S. exiguum 5.41 infective larvae per person (95% CI: 0.11 30.89 L3/person; MBR - July 1999 = 669.6). The ATPs were: (i) for S. oyapockense 18.25 infective larvae per person (95% CI: 0.45 105.53 L3/person; annual biting rates (ABR) = 17,802.0), (ii) for S. guianense 17.34 infective larvae per person (95% CI: 0.42 96.42 L3/person; ABR = 12,573.6), and for S. exiguum 10.50 infective larvae per person (95% CI: 0.22 59.95 L3/ person; ABR = 1,299.6) (Table III). In Toototobi, the prevalence of infection was 0.10% for S. oyapockense (n = 40 pools; with 38 negative pools - 95% CI: 0.00012 0.00370) and the prevalence of infective flies was 0.05% (n = 40 pools of 50 flies; with 39 negative pools - 95% CI: 0.00001 0.00282) (Table II). The highest MTP was in February 1999 (MBR = 5880) with 2.98 infective larvae per person (95% CI: 0.07 16.57 L3/person). The ATP of S. oyapockense was 13.59 infective larvae per person (95% CI: 0.34 75.66 L3/person; ABR = 26,851.6) (Table III). DISCUSSION The Brazilian Onchocerciasis Elimination Program has adopted the strategy suggested by OEPA that consists of semi-annual distribution of ivermectin (MectizanÔ) to at-risk communities to provide immediate clinical benefit and to reduce or eliminate transmission of O. volvulus in the Amazonian focus. To carry out a proper evaluation of the continuous chemotherapy provided by Funasa on transmission, two sentinel sites were chosen taking into consideration the endemic rates and the geographical location of the area Balawaú and Toototobi (Funasa 1995). The prevalence of infected flies is an important epidemiological tool to help decision makers to adopt new control strategies in a certain area. This could be accomplished by traditional entomological methods that are cumbersome and time consuming preventing the large scale application of the parasitological approach. Molecular methods based on PCR have proved to be sensitive and specific to address this necessity bringing the advantage of the possibility of performing the analysis in pools of insects, leading to a high throughput application. In the scenario of the aforementioned sentinel sites, where approximately 5000 simuliids were captured in one year, the only possibility of achieving reliable results was using the PCR method in pools of 50 insects. In addition, the prior dissection of the specimens generating three distinct anatomical pools head, thorax, and abdomen and further submission to the molecular method, allows the definition of infected and/or infective insects. Previous studies showed that PCR in si-muliids pools reveals prevalence of infected flies statistically similar to the rates obtained by traditional entomological dissection and parasite microscopic examination (Yamèogo et al. 1999, Rodríguez-Pérez et al. 1999). Py-Daniel (1994) compared the vector competency of S. oyapockense to S. guianense due to its (i) wide distribution in the Yanomami area (states of Amazonas and Roraima) and (ii) expressive number of biting specimens. These two factors contrast to the presence of the ciberal armadure that could hamper the transmission of O. volvulus. The potential of S. oyapockense in transmitting onchocerciasis in the area was observed in this study due to a positive PCR result for O. volvulus in pools collected in periods of high population density in Balawaú (470 specimens captured/MBR 5828) and Toototobi (525 specimens captured/MBR 5880). To define the prevalence of infected and infective flies, the vectors were individually dissected separating heads, thorax, and abdomens. The methodology described by Yamèogo et al. (1999) to separate the heads from the bodies, based on freezing of the specimens, was not efficient for S. oyapockense due to its reduced size. The rates found in this study showed that the prevalence of infection in simuliids collected in Balawaú decrease from 8.6 to 0.3% and in Toototobi from 4 to 0.1% in a four years interval of annual treatment, reinforcing that ivermectin administration does reduce O. volvulus transmission (Py-Daniel 1997, Guevara et al. 2003). According to Boatin et al. (1998), the treatment with ivermectin should be carried out in the beginning of the period of larger transmission corresponding to a larger impact. So, based on the results of this study, it is suggested that in Balawaú the treatment with ivermectin should be administered semi-annually, preferentially in January and July and in Toototobi in January and June. Considering that (i) the only definitive hosts that are capable of bearing O. volvulus are humans, (ii) the ivermectin distribution represents a long-term measure for the elimination of circulating microfilariae, (iii) the adult worms survive for a limit period of years, and (iv) insecticides cannot be efficiently applied in the breading simuliids sites in Yanomami areas, it seems that the control of the disease will only be accomplished with a competent surveillance program performed in parallel to the drug administration. This program has been developed in Brazil for the last 10 years and performance indicators are necessary to monitor the impact of the approach. The results obtained in this study, carrying out the molecular method for the first time in a Brazilian scenario, represent one of those indexes proving that the infection rates have decreased over time and should be routinely used in transmission areas under treatment with ivermectin, following the OEPA recommendation. ACKNOWLEDGEMENTS To Dr Tom Unnasch for introducing us to the O-150 detection and identification technique, and for providing the positive control (plasmid DNA pOVS134); to all the vector collectors of ONG CCPY and URIHI-Saúde Yanomami. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07034t1.jpg] [oc07034t2.jpg] [oc07034t3.jpg] [oc07034f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}