 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
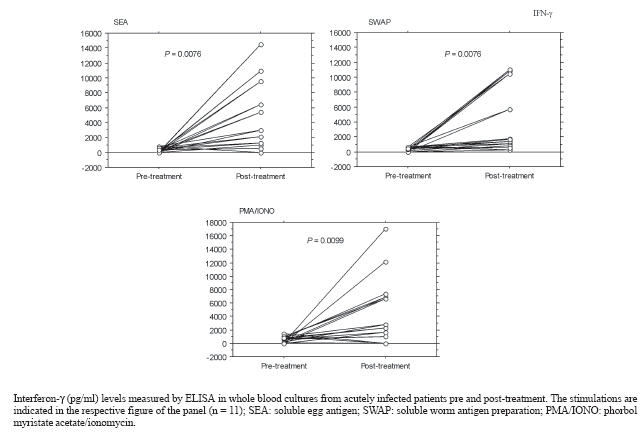
Memórias do Instituto Oswaldo Cruz, Vol. 102, No.2, March 2007, pp. 225-228 Treatment of human acute schistosomiasis with oxamniquine induces an increase in interferon-γ response to Schistosoma mansoni antigens Joelma R de Souza*/++, Clarice NL Morais, Marcílio L Aroucha**, Paulo JC Miranda****, Constança S Barbosa/+++, Ana Lúcia C Domingues**, Luiz B Carvalho Júnior*/***/+++, Frederico GC Abath /+++, Silvia ML Montenegro/+/+++ Centro de Pesquisas
Aggeu Magalhães-Fiocruz, Av. Prof. Moraes Rego s/no,
Cidade Universitária, 52020-020 Recife, PE, Brasil *Departamento
de Bioquímica **Hospital das Clínicas ***Departamento de Patologia
****Laboratório de Imunopatologia Keizo Asami, UFPE, Recife, PE, Brasil Financial support: CNPq, Capes, Fiocruz Received 17 August
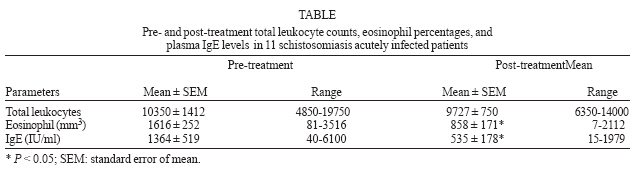
2006 Code Number: oc07039 Patients with acute schistosomiasis were studied before and after oxamniquine treatment. They had been exposed to cercariae 5 to 9 weeks before, and presented compatible clinical manifestations, eosinophilia, and high levels of total IgE. Interferon-g (IFN-γ) and interleukin-4 were measured by ELISA in whole blood samples under soluble egg antigen or soluble adult worm preparation stimulation. After treatment, the reduction of leukocytosis and eosinophilia were not significant, but total IgE levels decreased significantly, in contrast to IFN-γ levels that were significantly increased. The oxamniquine treatment of acute schistosomiasis patients is followed by an improvement of a Th1 response in vitro. If this response has a protective aspect is unknown, and some investigations need to be realized. Key words: Schistosoma mansoni - acute schistosomiasis - oxamniquine - cytokines In Brazil, schistosomiasis is historically considered to be a rural endemic disease. However, it is gradually expanding into some coastal regions of the state of Pernambuco, due to the exodus of rural workers to the urban areas, resulting in unusual outbreaks of acute schistosomiasis (Barbosa et al. 1998, 2000, 2001). Although immunological responses have been extensively examined in chronic patients, relatively few studies have been carried out with human acute schistosomiasis (Montenegro et al. 1999, Ribeiro de Jesus et al. 2002). In the present paper, patients selected from one of the rare outbreaks of acute schistosomiasis occurred in Brazil, were studied before and after oxamniquine treatment and the immunological findings reported. Eleven patients infected with acute schistosomiasis mansoni were selected in 2000, during an outbreak occurred in Porto de Galinhas beach, located in the southern coastal area of the state of Pernambuco, Brazil. They had been exposed to cercariae 5 to 9 weeks before, and presented fever, eosinophilia, cough, and, gastrointestinal symptoms. Informed consent was obtained from all patients, in line with the Brazilian ethical guidelines. The 11 subjects studied were 7 males and 4 females with ages ranging from 8 to 34 years (mean = 14 years). Physical, hematological, and stool examinations (according to Hoffman et al. (1934) and Katz et al. (1972) were performed in all patients. Detection of Schistosoma mansoni eggs was achieved in all acute patients studied. No other parasites were detected by stool examination. The parasite burden before treatment ranged from 24 to 1512 eggs/g of stool (mean = 220; SEM = 131). Blood samples (5 ml) were collected into heparin (10 U/ml) for leukocyte counts and cellular culture, before and 6 months after specific treatment with oxamniquine (15 mg/kg of body weight, single oral dose). Total blood leukocytes were stained with Turk's solution and counted in a Neubauer chamber. The relative and absolute eosinophil levels were determined in Giemsa stained blood smears. Whole blood without separation of peripheral blood mononuclear cells, were diluted in RPMI 1640 medium (1:3) plus penicilin (100 U/ml) and streptomicyn (100 μg/ml), and 0.7 ml were placed per well, as described previously (Montenegro et al. 1999). The cells were stimulated with soluble egg antigens (SEA, 20 μg/ml), soluble worm antigen preparations (SWAP, 50 μg/ml), and phorbol myristate acetate (PMA, 50 ng/ml) plus ionomycin (IONO, 1 μg/ml) as mitogens in a 48 well flat-bottom plates and incubated at 37oC in a 5% CO2 humidified atmosphere. Then, supernatants were collected after 96 h, quickly frozen, and stored at 70oC until use. Interferon-g (IFN-γ) and interleukin-4 (IL-4) were quantified by capture ELISA (enzyme linked immunosorbent assay) as described previously (Montenegro et al. 1999). Eleven healthy Brazilian individuals, living in non-endemic areas of Pernambuco, with negative stool examinations and no previous history of exposure to schistosoma infection were included as controls. Total IgE (E immunoglobulin) antibodies were measured in plasma by the ACCESS (Beckman Coulter) immunoassay, as described elsewhere (Addison et al. 1972). Differences between paired groups were assessed by the Wilcoxon signed rank test and correlation between variables was determined by calculating the Spearman's rank correlation coefficients. Statistical significance was considered at P < 0.05. Most of the reinfection studies indicate that schistosome-specific Th2 responses are associated with protective immunity after treatment of chronically infected patients (Butterworth 1998). In this context, it has been previously suggested that drug-dependent killing of worms in situ might boost protective immunity against reinfection by immunizing with antigens released from dying worms (Dunne et al. 1992, Webster et al. 1997, Woolhouse & Hagan 1999). More recently, it was shown that the immune responses induced by chemotherapy is heterogeneous, depending on the type of antigen used and time of analysis after treatment (Fitzsimmons et al. 2004, Joseph et al. 2004). However, most of the immunological studies after schistosomiasis treatment were undertaken in chronically infected patients living in endemic areas. We studied the effects of oxamniquine treatment in acutely infected patients that did not live in endemic areas and were selected during an outbreak of acute schistosomiasis in a coastal area of Brazil. Although the cytokine assays were not performed in a large sample, the present paper contributes to a better understanding of the immune responses after treatment, because acute schistosomiasis is rare in endemic areas. Our results demonstrated that after treatment the IFN-γ levels increased significantly in response to SEA (P = 0.0076), SWAP (P = 0.0076), and mitogens (P = 0.0099, Figure). Before treatment, the production of IFN-γ was similar to controls (data not shown). This is in line with studies that demonstrated increased IFN-γ production by T cell clone populations derived from treated acute patients (Contigli et al. 1999). Similarly, it was reported that patients with chronic disease living in endemic area showed increased levels of IFN-γ after treatment, in response to parasite antigens (Correa-Oliveira et al. 2000). Probably, the increase in T cell reactivity after chemotherapy can be explained by exposure of released antigens to the immune system following destruction of worms by chemotherapy (Araújo et al. 1996, Grogan et al. 1996, Brito et al. 2000, Correa-Oliveira et al. 2000). Nonetheless, the production of IFN-γ post-treatment did not correlate significantly with the parasite burden before treatment, possibly because of the small number of patients analyzed. It has been suggested that schistosome infection downmodulates the immune responses (Colley et al. 1986, Zwingenberger et al. 1989). Thus, alternatively, the increase in T cell reactivity after the treatment of infection, could be explained by termination of modulation and release from the repression of the immune response (Joseph et al. 2004). Although there are evidence for a role of IL-10 in the suppression of the Th1 response in late stages of the acute phase (16 weeks after exposure) of human schistosomiasis (Montenegro et al. 1999, Abath et al. 2006), it is not clear if this would occur at earlier stages of acute human schistosomiasis. On the other hand, there are reports that levels of IFN-γ induced by SEA and SWAP were not significantly changed by treatment in chronic patients from endemic areas 7 weeks after treatment with praziquantel (Joseph et al. 2004). Chronically infected patients treated with pra-ziquantel produced significantly more IL-4 and IL-5 in response to SWAP, although this was not observed in response to SEA (Joseph et al. 2004). In contrast, we could not demonstrate a significant increase in the production of IL-4 after treatment of acute schistosomiasis, as production of IL-4 in response to SEA or SWAP were similar to controls before and after chemotherapy (data not shown). The majority of studies on human immune responses highlighted the importance of humoral immunity based on IgE responses, having eosinophils as effector cells (Hagan et al. 1991). Herein we showed that acutely infected patients presented leukocytosis with eosinophilia and high levels of total IgE. Plasma IgE levels (Table) and eosinophil counts were significantly decreased after treatment (P = 0.0164 and 0.0208, respectively). Differently, studies in chronic patients demonstrated an increase in the levels of IgE antibodies against adult worm antigens after three months of treatment (Correa-Oliveira et al. 2000). In summary, treatment of acute schistosomiasis with oxamniquine is followed by an improvement of a Th1 response in vitro. If this response has a protective character is unknown and demands further investigations. ACKNOWLEDGEMENTS To Wlademir G de Melo for his technical assistance REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07039t1.jpg] [oc07039f1.jpg] |
| |||||||||

{kind=link}
{kind=link}