 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No.2, March 2007, pp. 233-235 What does determine gonad weight in the wild kissing bug Mepraia spinolai? Carezza Botto-Mahan+, Rodrigo Medel Departamento de
Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Santiago,
Chile Financial support: Fondecyt 3050033 (CBM) Received 15 September
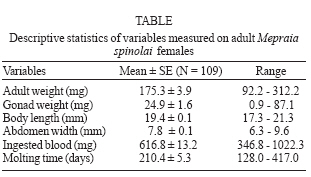
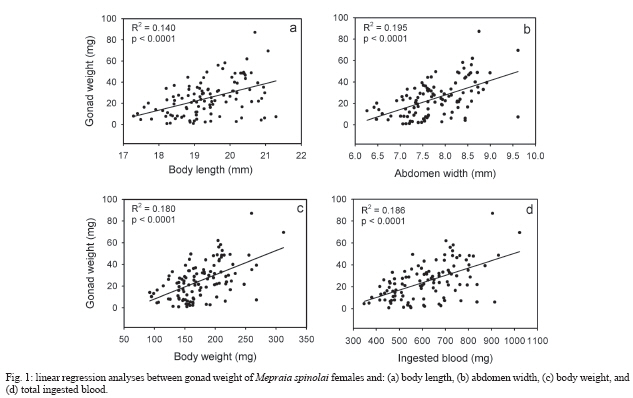
2006 Code Number: oc07041 Female fecundity increases with body size in a variety of insects, but it is unknown if this generalization applies for kissing bugs. In this study, we evaluate whether gonad weight in the bloodsucking insect Mepraia spinolai correlates with body size, or determined by nutrition or developmental time. We found that the investment on reproductive tissue correlates positively and significantly with body size and with the amount of ingested blood by female insects along their lifespan. Total molting time did not significantly affect gonad weight. We suggest that under optimal feeding conditions M. spinolai females could express their maximum reproductive potential. Key words: Triatominae - Reduviidae - reproductive investment Female fecundity has been shown to increase with body size in a variety of insects. Size and fecundity are influenced by genetic factors, development, and environmental conditions, however, under constant environmental conditions fecundity is positively correlated with female size (Honek 1993). Egg production increases with body size and this increase is often expected simply because larger females have more internal space for eggs. In addition, when female reproduction is based on energy reserves accumulated over long periods, large female size may confer fecundity advantage because the capacity for storing energy reserves increases more rapidly with body size than do metabolic costs (Andersson 1994). Unfortunately, no studies have experimentally examined the relationship between different measurements of body size (body length, abdomen width, and body weight) and reproductive tissue investment considering long-life nutrition and developmental time as potential variables affecting the fecundity of female insects. In this study, we evaluate whether gonad weight in the bloodsucking insect Mepraia spinolai is positively correlated with any component of body size, or determined by nutrition or total molting time. The kissing bug M. spinolai (Reduviidae; Triatominae) is one of the vector species responsible of Trypanosoma cruzi transmission in arid and semiarid Chile (Lent et al. 1994, Botto-Mahan et al. 2005). This strictly hematophagous and diurnal insect distributes between 18º and 34ºS and its main habitat includes stay grounds, bird nests, rock crevices, and caves (Lent & Wygodzinsky 1979). M. spinolai requires blood of vertebrates to complete its life cycle (Sagua et al. 2000, Canals et al. 2001). The development of this hemimetabolous insect includes an egg, five instar nymphs, and the adult. Often one full engorgement is sufficient for molting from one instar nymph to the next (Kollien & Schaub 2000). M. spinolai eggs were obtained from laboratory matings between adults collected at Las Chinchillas National Reserve (31º30'S, 71º06'W, IV Region, Chile; Botto-Mahan et al. 2002). Eggs were daily isolated from the mating jars, cleaned, and placed in sterile plastic containers. Each nymph was individually housed in a 3.2 cm x 3.6 cm clear plastic compartment of an 18-compartment box (11.4 cm x 20.5 cm). Each compartment was provided with sandy bottom and a folded piece of paper as refuge. All insects were reared under optimal growing conditions in a climatic chamber at 26ºC, 65-70% relative humidity, and 14:10 h light:dark regime (Ehrenfeld et al. 1998). Nymphs were fed 3-4 weeks after eclosion, using laboratory mice (C3H mouse strain, Central Breeding Ground, Facultad de Medicina, Universidad de Chile, Santiago, Chile). After each moult, nymphs were starved for 3 weeks and then fed on mice until adult emergence. Nymph weight was recorded before and after feeding. Molting was recorded on a daily basis and once adults emerged, sex was determined according to sexually dimorphic characters (Lent & Wygodzinsky 1979). Female nymphs (N = 109) reaching maturity were weighted and photographed with a digital camera (Sony Cybershot DSC-F717 5 MP) one day after molting to adult. Female length and width measurements were obtained from the digital photograph using UTHSCSA Image-Tool for Windows, version 2.01 (Wilcox et al. 2000). The body length was measured as the distance between the head and the last segment of the abdomen. Abdomen width was measured as the wider part of the abdomen (IV-V intersegmental suture). One month after maturity, bugs were dissected to extract and weight their fresh ovaries. Nutrition was quantified as the total weight of the blood ingested along insect ontogeny. All weight measurements were performed in an analytic scale (precision 0.1 mg). Molting time was calculated as the total number of days elapsed from the initial feeding as first instar nymph until adult emergence. To examine whether body size (i.e., body length, abdomen width, and body weight) nutrition and/or developmental time explain gonad weight of adult female bugs, we performed independent regression analyses. Bon-ferroni adjustment with a tablewide a-level = 0.05 was carried out to check for Type I error after performing multiple independent statistical analyses (Rice 1989). Variables were checked for homogeneity of variance by using the Fmax test. To obtain normality, variables were log-transformed when necessary (Sokal & Rohlf 1995). In M. spinolai, ovary weight represents on average (± SE) 13.81% (± 0.76%) of the total body weight. The Table summarizes descriptive statistics of the variables measured on adult female bugs. Our results showed that gonad weight positively and significantly correlates with all measurements of body size: body length, abdomen width, and body weight (Fig. 1a, b, c, respectively), and with the total amount of ingested blood during the bug ontogeny (Fig. 1d). On the other hand, ovary weight did not correlate with the total time needed to reach maturity (p = 0.469). All significant regressions kept significant after Bonferroni adjustment. These results indicate that body size and long-life nutrition accounted for an important fraction of the variance in gonad weight. In this study, we found that the investment on reproductive tissue mainly depends on body size components and the amount of ingested blood by M. spinolai females along their lifespan. As expected, gonad weight was higher for larger and better-nursed female insects. Hurd et al. (1995) suggested that female body size might affect reproduction in two ways. First, fecundity can be constrained by the number of ovarioles present in each ovary, which is often a function of body size. Second, body weight reduction may affect blood feeding and bloodmeal utilization for egg production. Our findings support the longstanding idea that body size and fecundity are positively correlated in insects (Honek 1993). Therefore, at least in female kissing bugs, body weight as well as linear measurements of body size would be reliable predictors of reproductive investment. In addition, our results indicate that long-life nutrition should be a variable to be considered when examining reproductive investment and could be an indicator of environmental conditions during insect development. The amount of ingested blood may have also an indirect effect on the investment in reproductive tissue through an increase in body size of female bugs, probably due to correlations between body size and many body organs. On the other hand, developmental time had a slightly negative but not significant effect on gonad weight. If we assume that organisms have limited resources for reproduction, and that resource allocation to defense, growth and maintenance compromise resource allocation to reproduction (Forbes 1993, Perrin et al. 1996), the observed tendency could be explained because female bugs needing more time to reach maturity would divert more energy to somatic maintenance and survival instead of reproduction. Overall, the results of our study suggest that under optimal feeding conditions M. spinolai female bugs could express their maximum reproductive potential through a direct effect of nutrition on gonad weight and probably indirectly through body size. ACKNOWLEDGMENTS To Dr Madeleine Lamborot for helping with the gonad extraction technique and CG Ossa for technical support. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07041f1.jpg] [oc07041t1.jpg] |
| |||||||||

{kind=link}
{kind=link}