 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No.3, June 2007, pp. 329-334 Antimalarial drugs disrupt ion homeostasis in malarial parasites Marcos L Gazarini, Carlos AO Sigolo, Regina P Markus, Andrew P Thomas*, Célia RS Garcia/+ Departamento
de Fisiologia, Instituto de Biociências, Universidade de São
Paulo, Rua do Matão, travessa 14, no 321,
05508-900 São Paulo, SP, Brasil *Department of
Pharmacology and Physiology, University of Medicine and Dentistry
of New Jersey, Newark, New Jersey, US Financial support: Fapesp, CNPq Receive
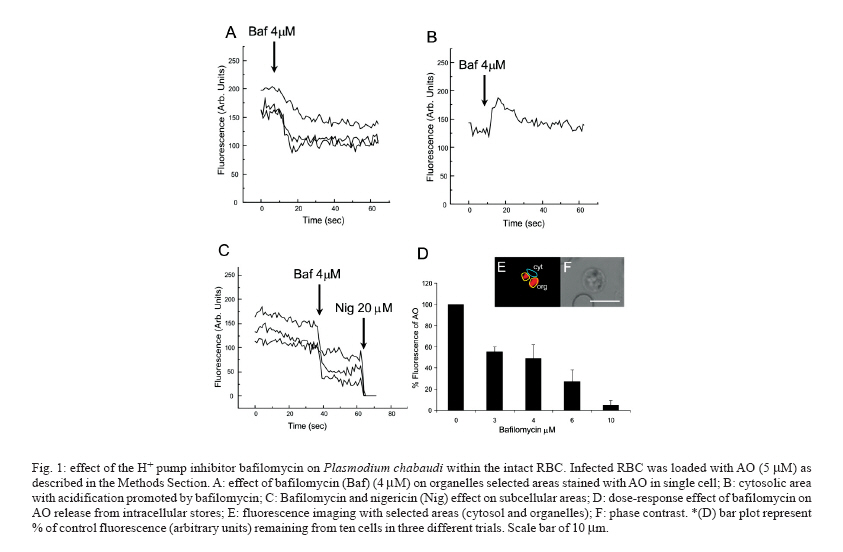
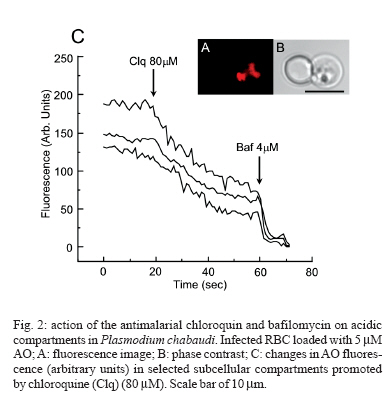
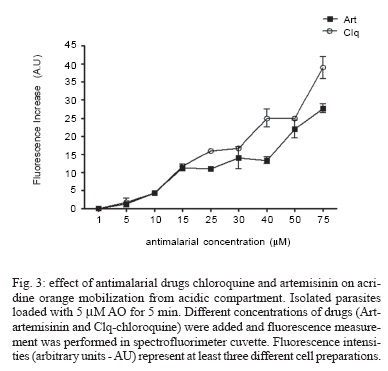
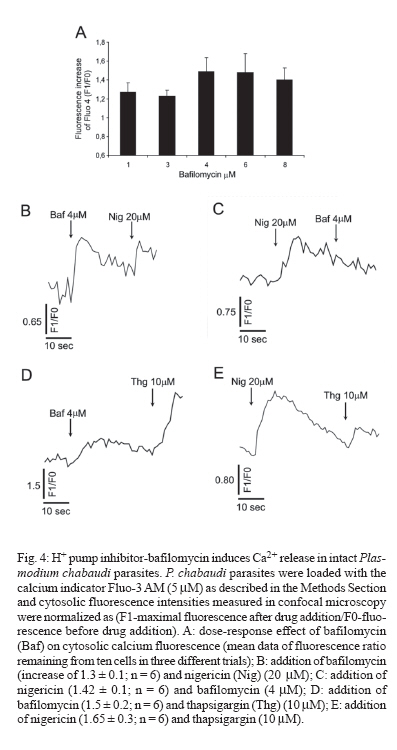
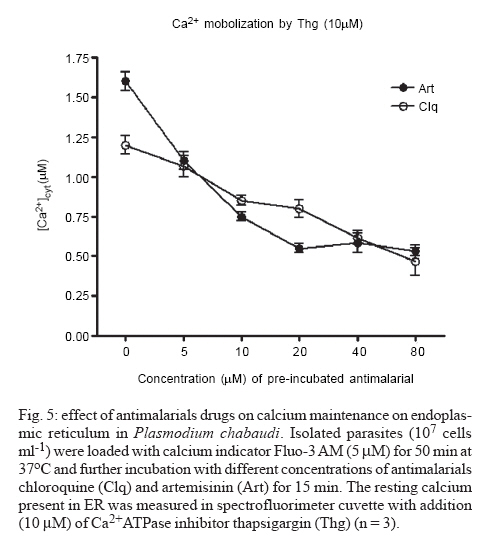
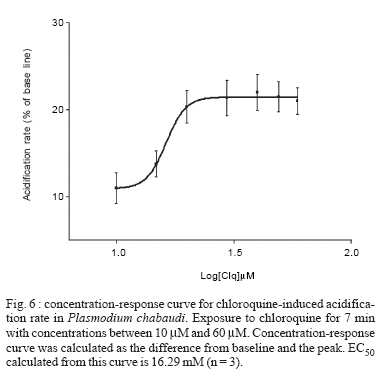
8 February 2007 Code Number: oc07055 Plasmodium chabaudi malaria parasite organelles are major elements for ion homeostasis and cellular signaling and also target for antimalarial drugs. By using confocal imaging of intraerythrocytic parasites we demonstrated that the dye acridine orange (AO) is accumulated into P. chabaudi subcellular compartments. The AO could be released from the parasite organelles by collapsing the pH gradient with the K+/H+ ionophore nigericin (20 μM), or by inhibiting the H+-pump with bafilomycin (4 μM). Similarly, in isolated parasites loaded with calcium indicator Fluo 3-AM, bafilomycin caused calcium mobilization of the acidic calcium pool that could also be release with nigericin. Interestingly after complete release of the acidic compartments, addition of thapsigargin at 10 μM was still effective in releasing parasite intracellular calcium stores in parasites at trophozoite stage. The addition of antimalarial drugs chloroquine and artemisinin resulted in AO release from acidic compartments and also affected maintenance of calcium in ER store by using different drug concentrations. Key words: malaria - Plasmodium chabaudi - calcium - chloroquine - artemisinin - acidic compartment During the course of its intraerythocytic life, Plasmodium causes well documented biochemical and morphological changes in both pathogen and host cell (Howard 1982, Coppel 1986, Garcia et al. 1997, Kirk 2001). Acidic compartments are organelles present in Plasmodium and several trypanosomatids (Docampo & Moreno 2001). For Plasmodium, it has been suggested that these organelles are the food vacuole in which hemoglobin is digested and antimalarial drugs such as chloroquine are accumulated (Yayon et al. 1984). Chloroquine is though to act by preventing the monomeric hemin from forming the inert and insoluble polymer hemozoin which is derived from digestion of hemoglobin in the acidic vacuole (Slater & Ceram 1992). Defining the sites of chloroquine action in Plasmodium, as well as determining how it is related to proton activity in the acidic compartment is of major importance in the physiology of these parasites.It has been reported that the antimalarial artemisinin act on PfATP6 (Ca2+ ATPase) in P. falciparum, thus affecting Plasmodium Ca2+ homeostasis (Uhlemann et al. 2005). Regulation of internal pH is important to parasite survival as this sets the environment for intracellular enzymes to function. It has been reported that P. fal-ciparum and P. chabaudi extrude H+ to the extracellular medium by a Na+ independent mechanism (Saliba & Kirk 1999). Other mechanisms for pH regulation described in malaria parasites may involve the V-H+-ATPase and the V-H+-PPase in acidocalcisomes (Mar-chesini et al. 2000, McIntosh & Vaidya 2002, Saliba et al. 2003, Biagini et al. 2003). We have reported the existence of Ca2+ pools in the acidic compartment of rodent as well as human malaria parasites as a possible mechanism for the regulation of calcium concentration in these parasites (Garcia et al. 1998, Passos & Garcia 1998). Calcium homeostasis in malaria parasites has been extensively studied by several authors (Tanabe et al. 1982, Krungkrai & Yuthavong 1983, Scheibel et al. 1987, Wasserman et al. 1990, Adovelande et al. 1993, Garcia et al. 1997, 1998, Gazarini et al. 2003, Gazarini & Garcia 2004, Beraldo et al, 2005, Budu et al. 2007). Interestingly, in P. chabaudi, the second messenger IP3, mobilizes Ca2+ not only from the classical ER-like pool but also from an acidic compartment, implying that these parasites utilize a calcium-mediated cell signaling mechanism similar to higher organisms (Passos & Garcia 1998). Nevertheless, we still have no clear picture of signaling events and the role of the acidic compartment in malaria parasite signaling, or its significance in the therapeutic actions of antimalarial drugs such as chloroquine (Yayon et al. 1984, Dzekunov et al. 2000). Ca2+ and pH changes are crucial for parasite enzyme activity such as proteases (Na et al. 2004, Farias et al. 2005). In this context, understanding the mechanisms utilized by Plasmodium for the maintenance of its ion homeostasis is crucial. In the present work, we have studied the physiological mechanisms by which H+ and Ca2+ activity are related in the acidic compartments of P. chabaudi, and how these are affected by the antimalarials chloroquine and artemisinin. Unraveling the mechanisms by which these organelles contribute to cell function may be important for developing alternative strategies for therapeutic approaches to malaria. MATERIALS AND METHODS P. chabaudi parasites - Synchronous parasitemia by P. chabaudi (clone AJ) was maintained in female mice (Balb/c) by transfer of infection. Parasitemia was about 50%. After collecting the blood, cells were washed with PBS (137 mM NaCl, 2.7 mM KCl, 4.3 mM Na2HPO4, 1.4 mM NaH2PO4 and 1 mM CaCl2) and leucocytes and platelets were removed by a powdered cellulose column. Parasitemia was determined from Giemsa-stained thin films. Parasite isolation - The infected RBCs were then washed twice by centrifugation at 1500 g for 7 min in PBS. Infected RBC (107 ml-1) was lysed in PBS with 60 µg.ml-1 saponin. RBC membranes were removed by centrifugation (9000 g for 10 min at 4oC), the parasites were washed twice in MOPS buffer (116 mM NaCl, 5.4 mM KCl, 0.8 mM MgSO4, 5.5 mM D-glucose, 50 mM MOPS, and 2 mM CaCl2, pH 7.2) and dispensed in the same buffer at room temperature (Passos & Garcia 1998). Confocal microscopy - In vitro parasite-infected RBCs (106 cells ml-1) were loaded with acridine orange (AO) (5 μM) for 5 min at room temperature and washed once with MOPS buffer as described above. Cells were resuspended in the same buffer and placed in a microscopy chamber. The data acquisition was performed with Zeiss confocal microscope (LSM 510) using the LSM 510 software, version 2.5; excitation 488 nm (Argon laser) and emission collected with a bandpass filter of 505-550 nm (Fluo3-AM). Calcium measurements were performed with isolated parasites incubated with Fluo-3 AM (5 µM) for 50 min at 37oC and placed in microscopy chambers at room temperature. The configurations for image acquisition were the EX 488 nm and Em > 560 nm for AO measurements. Software-based analysis allowed fluorescence imaging in the whole field of view or in a selected cell as a function of time. This was accomplished by defining areas of interest on a given image frame and requesting the software to construct a graphical representation of intensity against time. Experiments were carried out with at least three different cell preparations. Traces represent typical single cell responses unless indicated otherwise. Spectrofluorimetric measurement - Isolated parasites (107 cells ml-1) were incubated with MOPS buffer in 2 ml cuvette. The measurements were performed in a Shimadzu RF-5301 PC spectrofluorimeter at 37oC and dyes fluorescence (Fluo-3 AM and AO) was measured continuously after the drug addition. Parasites were incubated with calcium indicator Fluo-3 AM (5μM) for 50 min at 37oC in MOPS buffer containing 2 mM CaCl2 and probenecid (40 μM) or AO (5 μM) incubated for 5 min at 37oC in the same buffer. The dyes parameters are excitation 490 nm and emission collected at 530 nm for Fluo-3 AM and AO fluorescence measurement was obtained with Ex-495 nm/Em-590 nm. Extracellular H+ levels - The effect of chloroquine on H+ mobilization in isolated parasites was recorded using a Cytosensor microphysiometer (Mol Devices, US), a silicon-based biosensor system. This instrument continuously monitors the extracellular pH surrounding cells in culture, and reports cell activation by measuring the increases in extracellular acidification rate occurring in response to agonist stimulation (McConnell et al. 1992). The isolated parasites was resuspended in DMEM (GIBCO) plus 30% agarose to improve parasites adherence onto the polycarbonate membrane of the transwell capsules (3 μm pore size) at a density of 2.5 x 105 cells. RESULTS We have investigated the dynamics of solute exchange in the acidic compartments of isolated and intact blood-stage malaria parasite, P. chabaudi. To study intracellular pH changes, we loaded infected red blood cells at trophozoite stage (P. chabaudi) with AO. Dye mobilization through compartments was measured by selecting areas of interest (cytosol and acid compartment) by confocal imaging (Fig. 1). Addition of H+-ATPase inhibitor bafilomycin (4 mM) to the cells led to alkalinization of acidic compartments as indicated by the partial release of accumulated AO (Fig. 1A), and simultaneously increases the cytosol fluorescence (Fig. 1B). The remaining AO could be released by addition of the K+/H+ ionophore nigericin to the cells (20 μM), which totally released the compartmentalized AO fluorescence (Fig. 1C). Dose-dependent addition of bafilomycin promotes the depletion of AO stores. A remarkable (over 90%) release of AO fluorescence occurs with addition of bafilomycin at 10 μM (Fig. 1D). The effect of the antimalarial chloroquine on AO accumulation in subcellular compartments (Fig. 2A) in infected erythrocyte (Fig. 2B) shows that chloroquine (80 μM) promotes extrusion of AO from the acidic compartments, presumably as a result of alkalinization. The subsequent addition of H+-ATPase inhibitor bafilomycin (4 μM) caused complete depletion of the residual AO from the acidic compartments (Fig. 2C). Taken together, the data of Figs. 1 and 2 suggest that P. chabaudi parasites display a bafilomycin-sensitive acidic compartment that can be affected in a dose-dependent manner and is also a target for chloroquine. By using spectrofluorimeter measurements and isolated parasites (107 cells) at trophozoite stage previously incubated with AO (5 μM) for 5 min/37oC we showned that AO could be released in a dose-dependent manner by addition of artemisinin and chloroquine (Fig. 3). We have previously demonstrated that the H+-pump inhibitor 7-chloro-4-nitrobenz-2-oxa-1,3-diazole (NBD-Cl) caused release of Ca2+ in Fluo-3 loaded P. falciparum parasites (Garcia et al. 1998). We therefore investigated the effects of the more selective H+-pump inhibitor, bafilomycin on Ca2+ homeostasis. We measured the fluorescence changes in isolated parasites (trophozoite stage) loaded with the calcium probe Fluo-3 AM (5 μM) and placed to microscopy microplates for confocal measurements, in response to effectors of the acidic compartments. Using different concentration of bafilomycin, we observed that bafilomycin-sensitive Ca2+ pool is completely discharged at 4 μM (Fig. 4A). However subsequent addition of nigericin (20 μM) after bafilomycin treatment (4 μM) reveals a subsequent Ca2+ rise (Fig. 4B). As nigericin was able to release the entire Ca2+ pool by itself further addition of bafilomycin had no effect on Ca2+ release (Fig. 4C). Interestingly, neither bafi-lomycin (Fig. 4D) or nigericin (Fig. 4E) was able to deplete ER Ca2+ pool, as a subsequent addition of thapsigargin (Thg) promoted a second calcium rise. To investigate the effect of the antimalarials drugs chloroquine and artemisinin in calcium homeostasis we carried out experiments with isolated parasites loaded with calcium indicator Fluo-3 AM (5 μM). Fig. 5 shows spectrofluorimetric measurements of calcium release by Thg (10 μM) with parasites previously incubated (15 min at 37oC) with different concentrations of chloroquine and artemisinin (0-80 μM) in a high calcium extracellular medium (2 mM). The results show that previous parasite incubation with chloroquine or artemisinin leads to a dose-dependent reduction of total amount of calcium release by thapsigargin. When parasites are exposed to chloroquine, the cytosolic pH decrease as a result of extrusion of H+ from acidic compartments and the parasites (P. falciparum) try to restore the cytosolic pH level using plasma membrane mechanisms (Saliba & Kirk 1999, Marchesini et al. 2000, Saliba et al. 2003). By using microphysiometry we showed that addition of chloroquine to P. chabaudi at the trophozoite stage led to a dose-dependent increase in the extracellular acidification rate (Fig. 6). The mean 50% maximal effective concentration (EC50) for three experiments was 16.29 ± 0.4 μM (n = 3) of chloroquine. This data reveal an activation of plasma membrane mechanism of H+ extrusion under chloroquine treatment. DISCUSSION The antimalarial drug chloroquine is thought to accumulate in acidic compartments, although its mechanisms of action are controversial (Slater & Cerami 1992, Dorn et al. 1995, Waller et al. 2003). The resistance to chloroquine observed in P. falciparum is dependent to mutations in specific transporter (PfCRT), by using heterologous expression Reeves et al. (2006) observed a increase of lysosomes acidification in mammalian cells expressing PfCRT, providing new information about acidification process and chloroquine resistance. We have previously shown that the acidic pool also functions as a Ca2+ store in permeabilized malaria parasites and that chloroquine causes Ca2+ release from this store (Passos & Garcia 1998). In the present study, intact parasites were labeled with the calcium indicator, Fluo-3 AM to measure parasite cytosolic Ca2+ mobilization, and we also investigated the acidic pools in intact parasites within the RBC, using confocal microscopy. The data in the present work indicate the existence of mechanisms for H+ homeostasis in P. chabaudi parasites. The acidic vacuole in malaria parasite is the site of hemoglobin digestion and it is believed of drug action. In the rodent malaria parasite P. chabaudi the second messenger IP3 is known to induced-Ca2+ mobilization from this compartment (Passos & Garcia 1998). These findings are of major significance, because they raise the possibility that chloroquine and artemisinin action in malaria parasites may involve alterations in ion homeostasis. This is especially important in view of our work showing that IP3-dependent Ca2+ signaling is involved in the progression of the malarial parasite cell cycle (Hotta et al. 2000, Gazarini et al. 2003, Beraldo et al. 2005). By using confocal microscopy and analyzing P. chabaudi loaded with AO within the intact RBC, we showed a H+ mobilization, corresponding to a fluorescence decrease when V-type H+-ATPase was inhibited by bafilomycin (Fig. 1). Simultaneously, parasite cytosol fluorescence was transiently increased as the AO was released, thus indicating that cellular mechanisms were operating to restore cytosolic pH (Fig. 1B). Bafilomycin completely discharge the AO pool at 10 μM (Fig. 1D) while Ca2+ release reach the maximal values at addition of 4 μM of H+ pump inhibitor (Fig. 4A). The interaction of antimalarial artemisinin in P. falciparum was reported to occur through inhibition of PfATP6, a SERCA-type ATPase (Uhlemann et al. 2005). Our data shows the ability of both antimalarial in modifying the calcium and proton dynamics in P. chabaudi internal stores (Fig. 5). We also addressed the metabolic response evoked by chloroquine with cytosensor microphysiometer (Fig. 6). These results showed that chloroquine stimulated a H+ extrusion response with an EC50 of 16 μM in P. chabaudi, using plasma membrane mechanisms to recover the intracellular pH (Saliba & Kirk 1999). Our data provide new information of acidic organelle physiology, the high concentration of bafilomycin required for complete reversion fluorescence from AO, suggests a low density of H+ pumps or susceptibility to the drug. However chloroquine action promoted a more extensively effect on parasite ion homeostasis than the specific H+ pumps inhibitor bafilomycin. The micromolar concentration of antimalarial drugs required in our assay may represent the cell physiology differences between the Plasmodium species and we are search for extension of organelles capacity in ion maintenance. The fact that Ca2+ antagonists block malaria parasite development is well known. In addition, reversal of chloroquine resistance in vitro by several Ca2+ antagonists such as verapamil has also been extensively reported (Deloron et al. 1991, Adovelande et al. 1998). We have demonstrated that host melatonin activates parasite calcium signaling via the second messenger IP3, which appears to be important in the proliferation and maturation of the intraerythrocytic malaria parasites (Hotta et al. 2000). More recently, we also verify calcium mobilization with AFMK, a product of melatonin degradation in Plasmodium-infected erythrocytes (Budu et al. 2006). Finally, although the molecular mechanism of chloroquine and artemisinin action is not fully understood, its affect on calcium and proton activity indicates the importance in elucidate calcium homeostasis and signaling mechanisms on malarial parasites. ACKNOWLEDGEMENTS To Fapesp for funding CRSG and RPM. MLG received fellowship from Fapesp and CAS from CNPq. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07055f6.jpg] [oc07055f5.jpg] [oc07055f1.jpg] [oc07055f2.jpg] [oc07055f4.jpg] [oc07055f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}