 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No.3, June 2007, pp. 421-424 Merozoite surface protein 2 allelic variation influences the specific antibody response during acute malaria in individuals from a Brazilian endemic area Selma Sallenave-Sales, Clarissa Perez Faria, Mariano Gustavo Zalis*, Cláudio Tadeu Daniel-Ribeiro, Maria de Fátima Ferreira-da-Cruz/+ Departamento
de Imunologia, WHO Collaborating Center for Research and Training
in the Immunology of Parasitic Diseases, Instituto Oswaldo Cruz
-Fiocruz, Av. Brasil 4365, 21040-900 Rio de Janeiro, RJ, Brasil
*Departamento de Medicina Preventiva, Hospital Universitário
Clementino Fraga, UFRJ, Rio de Janeiro, RJ, Brasil Financial support: IOC-Fiocruz, CNPq Received
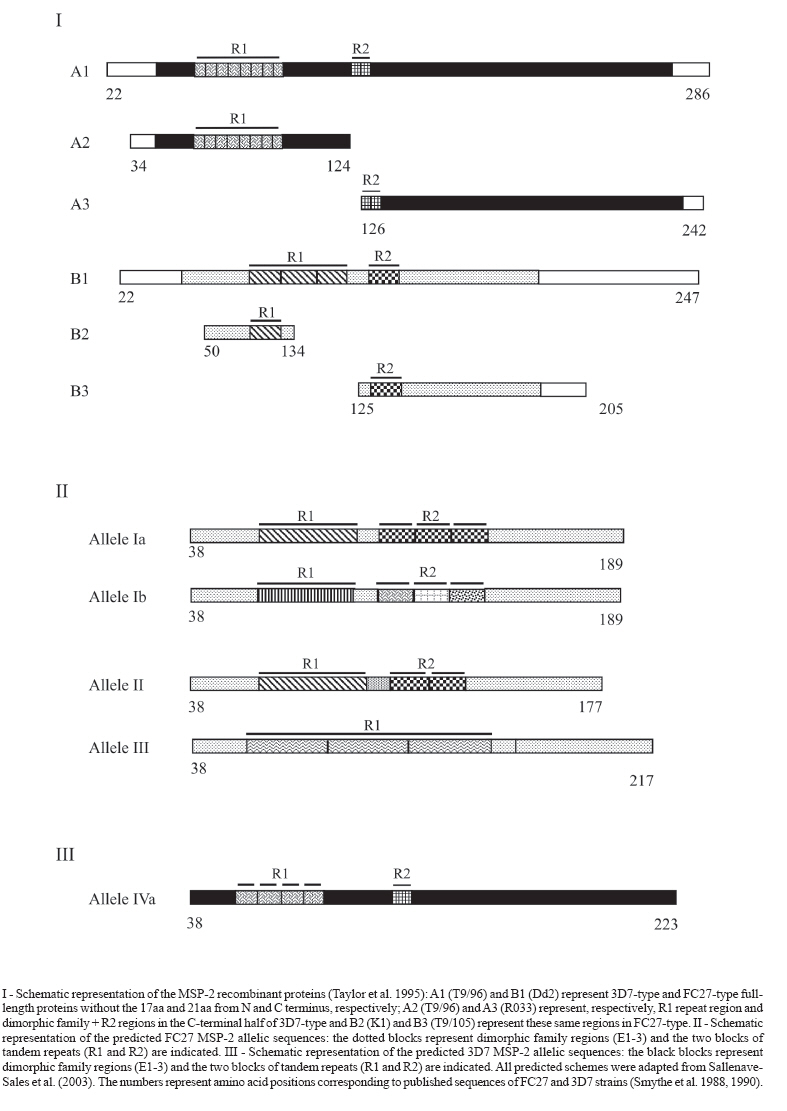
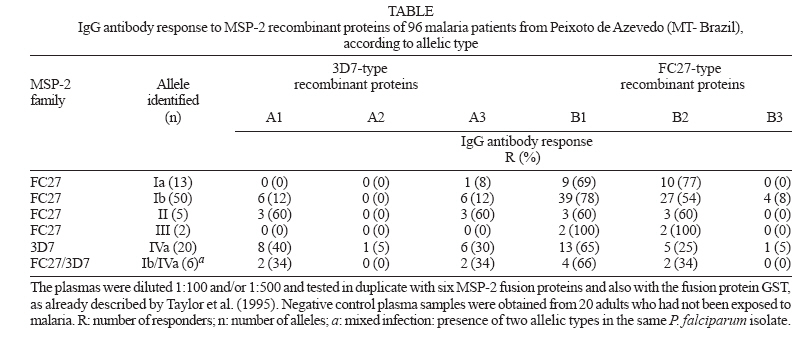
2 March 2007 Code Number: oc07067 The antibody response to Plasmodium falciparum parasites of naturally infected population is critical to elucidate the role of polymorphic alleles in malaria. Thus, we evaluated the impact of antigenic diversity of repetitive and family dimorphic domains of the merozoite surface protein 2 (MSP-2) on immune response of 96 individuals living in Peixoto de Azevedo (MT-Brazil), by ELISA using recombinant MSP-2 proteins. The majority of these individuals were carrying FC27-type infections. IgG antibody responses were predominantly directed to FC27 parasites and were correlated to the extension of polymorphism presented by each MSP-2 region. This finding demonstrated the impact of the genetic polymorphism on antibody response and therefore, its importance on malaria vaccine efficacy. Key words: Plasmodium falciparum - merozoite surface protein-2 - antibody response - antigenic polymorphism - malaria It has been suggested that the genetic variability of the parasite could allow its evasion from immune responses induced by previous infections with parasites expressing variant antigenic forms. Genetic modifications in antigenic epitopes could induce conformational changes in the polypeptide structure resulting in the lack of putative reactive sites at the protein surface, as well as in different presentation of these sites to the immune system (Ranford-Cartwright et al. 1996, Tonhosolo et al. 2001, Kanunfre et al. 2003). Consequently, studies on the relationship of genetic diversity of B epitopes in parasite surface antigens and its impact on the recognition of antibodies generated during natural infections should be carried out for designing immunization strategies. The genetic polymorphism of the merozoite surface protein 2 (MSP-2) of Plasmodium falciparum has been studied due to its applicability as a target for a malaria vaccine development and its putative association with antibody-mediated protection (Taylor et al. 1995, Genton et al. 2003, Scopel et al. 2006). The MSP-2 allelic families known as ICI/3D7 and FC27 corresponding, respectively to two antigenically distinct serogroups: A and B, which are frequently and strongly recognized by antibodies from malaria-exposed individuals (Fenton et al. 1990, Ekala et al. 2002). These antibodies are preferably directed against the central variable region of the molecule, which is composed by tandem repeats blocks (R1 and R2) flanked by family dimorphic blocks. The FC27 family has shown different numbers of the relatively conserved R1 (32-mer) and R2 (12-mer) repetitive regions. The R1 repeats of 3D7 family, characterized by shorter sequence repeats of 3 to 10 residues, can vary considerably although they are usually glycine, serine and alanine rich, while the R2 region is a well conserved repeat rich in the amino acid threonine. It has been suggested that naturally immunodominant epitopes are encoded within the repetitive sequences of the molecule (Smythe et al.1990). Our group previously identified (Sallenave-Sales et al. 2003) polymorphic alleles from both MSP-2 families in P. falciparum isolates collected during a drug surveillance carried out between May 1995 and January 1996 in Peixoto de Azevedo village (state of Mato Grosso, Brazil). The village population consisted mainly in migrant individuals exposed during all the year to the risk of malaria infection. At time of blood collection, the village had the highest malaria transmission rates in Brazil with an annual parasitic incidence (positive slides/1000 inhabitants) of 284.8 and 263.1 in 1995 and 1996, respectively. After signed consent, venous blood samples were obtained, before treatment, from 96 adults between 20 and 60 years of age with uncomplicated malaria and MSP-2 alleles were typed by SSCP-PCR and sequencing analysis. Here, we intend to analize the impact of such antigenic polymorphism on patient antibody responses. For this purpose, we tested plasma from these 96 patients against six recombinant proteins representing the R1 units repeats and family dimorphic regions of MSP-2 families, by ELISA. Three MSP-2 recombinant proteins were tested to each one of serogroup (A and B): A1, A2, A3 (ICI/3D7) and B1, B2, and B3 (FC27) (Figure). A1 and B1 represent full-length proteins without the 21aa and 17aa from N and C terminus, respectively; A2 and B2 represent the R1 repeat region and A3 and B3 represent dimorphic family regions. These proteins were kindly provided by Dr Jana McBride group and the ELISA were performed as already described (Taylor et al. 1995). The frequency of individuals positive to at least one MSP-2 protein was 81% (data not shown). The IgG antibody prevalence to each of the six recombinant MSP-2 proteins is shown in the Table. As expected, we observed that recombinant proteins of serogroup B (FC27-types) were better recognized than serogroup A (3D7-types), since FC27 alleles were the predominant type detected on P. falciparum parasites infecting these individuals. Similar results using another set of MSP-2 recombinant proteins were reported by Kanunfre et al. (2003) studying malaria patients from the same village - Peixoto de Azevedo contrasting to data reported by Taylor et al. (1995) that even using the same recombinant proteins, observed a predominance of 3D7 antibodies (serogroup A) in a comparable analysis conducted in Gambia. We also detected a significant number of individuals responders to B1 (central variable region) and B2 (R1 region) recombinant proteins. Near 60% (44 out of 76) of the individuals that carried parasites type FC27 had antibodies to the first repetitive region of molecule (B2) that is relatively conserved even in geographically distinct plasmodial isolates. Interestingly, the two single individuals infected with allele III had antibodies to B2 recombinant protein, suggesting that the presence of three units at R1 region of this allelic type should contribute to the antigenicity of this recombinant protein (Figure). In contrast, the total deletion of R2 region in allele III sequence could be responsible for the lack of reactivity sites at the surface of the molecule (Figure). In fact, we observed that individuals carrying parasites with allele III do not respond to B3 recombinant protein which represents the dimorphic family region of FC27 type including R2 region. The non response or a low antibody response to MSP-2 recombinant protein containing one or no 12-mer repeats (units) in R2 region may indicate that such variation in the number of tandem repeats could allow the parasite to avoid high avidity antibody binding and it was already reported (Ranford-Cartwright et al. 1996). Curiously, we did not detected antibody response in individuals infected with parasites carrying alleles Ia or II, that shared two identical repeats in R2 region. In addition, only 8% of individuals infected with allele Ib type parasites recognized B3 recombinant protein, possibly due to sequence diversity between Brazilian MSP-2 genotypes and the reference strain (T9/105) that originate the recombinant protein (Figure). Contrasting to FC27-type recombinant proteins, only 20% (19 out of 96) of studied individuals presented antibodies to 3D7-type ones (serogroup A) (Table). The frequency of antibody response to A1 (central variable region) and A3 (dimorphic region including R2 repeats) were the same but only one patient had antibodies to the first repetitive region of molecule (A2). As observed in the case of B3 recombinant protein, low antibody recognition to A2 was detected in individuals carrying allele IVa, or carrying this allele in association to the allele FC27 Ib (Table). Once again this result could be related to sequence polymorphism of Brazilian MSP-2 genotypes in relation to the reference strain (T9/96) that originate the recombinant protein since amino acid sequence of the R1 repeats from different isolates could vary extensively (Figure) (Ferreira & Hartl 2007). In summary, the antibody response to B1 and B2 (FC27-type) and A1 and A3 (3D7-type) recombinant proteins may be associated to the extension of polymorphism presented by each MSP-2 region, since we found an inverse relationship between such polymorphism and the frequency of immune response. In respect to B3 recombinant protein, the variable number and sequence diversity of R2 repetitive units would be responsible for the low antibody reaction against this recombinant protein. Similarly, the high variability in length, sequence and copy number of R1 region, could explain the poor antibody reaction with the D2 recombinant protein. Finally, we conclude that the genetic polymorphism of MSP-2 protein in P. falciparum parasites could influence the development of specific antibody immune response in Brazilian populations. Analysis of the antibody response using recombinant proteins representing MSP-2 allelic types found in Peixoto de Azevedo village will done to better understanding the immune response to MSP-2 alleles. ACKNOWLEDGEMENTS To Dr Jana McBride for providing MSP-2 recombinant proteins. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07067t1.jpg] [oc07067f1.jpg] |
| |||||||||

{kind=link}
{kind=link}