 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No. 4, 2007, pp. 509-515 Cloning and characterization of a V-ATPase subunit C from the American visceral leishmaniasis vector Lutzomyia longipalpis modulated during development and blood ingestion JM Ramalho-Ortigão, AN Pitaluga, EL Telleria, C Marques, AA Souza*, YM Traub-Cseko/+ Laboratório de Biologia Molecular de Tripanossomatídios e Flebotomíneos, Departamento de Bioquímica e Biologia Molecular, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil *Instituto Evandro Chagas, Seção de Parasitologia, Belém, PA, Brasil Financial support: Papes IV-Fiocruz, IOC-Fiocruz +Corresponding author: ytraub@ioc.fiocruz.br JMR-O current address: Department of Biological Sciences, University of Notre Dame, 243 Galvin Life Sciences Center, Notre Dame, IN 46556, US Received
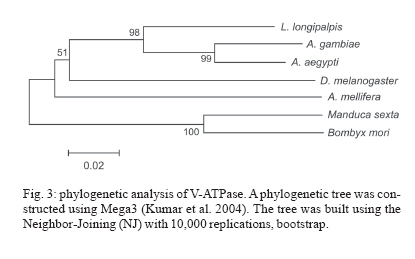
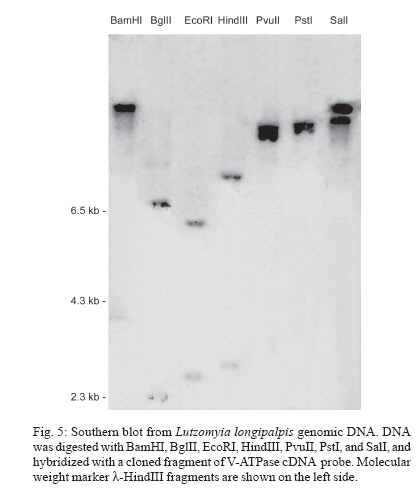
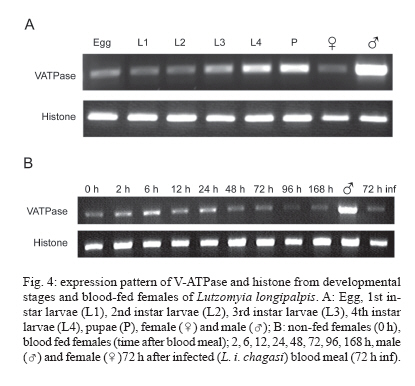
8 December 2006 Code Number: oc07080 Visceral leishmaniasis (VL) is a serious tropical disease that affects approximately 500 thousand people worldwide every year. In the Americas, VL is caused by the parasite Leishmania (Leishmania) infantum chagasi mainly transmitted by the bite of the sand fly vector Lutzomyia longipalpis. Despite recent advances in the study of interaction between Leishmania and sand flies, very little is known about sand fly protein expression profiles. Understanding how the expression of proteins may be affected by blood feeding and/or presence of parasite in the vector's midgut might allow us to devise new strategies for controlling the spread of leishmaniasis. In this work, we report the characterization of a vacuolar ATPase subunit C from L. longipalpis by screening of a midgut cDNA library with a 220 bp fragment identified by means of differential display reverse transcriptase-polymerase chain reaction analysis. The expression of the gene varies along insect development and is upregulated in males and bloodfed L. longipalpis, compared to unfed flies. Key words: Lutzomyia longipalpis - V-ATPase - metacyclogenesis - differential expression Leishmaniasis is an important public health problem in the whole world, with approximately 350 million people living in risk areas, 12 million people infected, and 2 to 3 million new cases every year. More than 90% of all cases of visceral leishmaniasis (VL) in the New World are in Brazil, with recent epidemic outbreaks even in urban areas (Antunes-Uchoa et al. 2001, Gontijo et al. 2002, Lainson & Rangel 2005). Lutzomyia longi-palpis is the principal vector for Leishmania infantum chagasi-caused VL (Bauzer et al. 2007). Molecular studies of disease transmitting vectors have led to the discovery of several molecules that hold promise in the development of new approaches to fight pathogen transmission. Great advances have been obtained mostly in malaria transmitting mosquitoes: the complete genome of Anopheles gambiae was published (Holt et al. 2002), and transgenic insects were obtained (reviewed in Marrelli et al. 2006). Unfortunately, however, the same cannot be said for sand flies, where similar types of studies are still incipient. Such a lack of information is even more impressive in regards to the American visceral leishmaniasis vector, L. longipalpis. Apart from the characterization of salivary gland genes (Charlab et al. 1999, Anderson et al. 2006), some recent work has focused on the sequencing of ESTs (Ramalho-Ortigão et al. 2001, Dillon et al. 2006) and the identification of a chitinase potentially involved in the biogenesis of the peritrophic matrix (Ramalho-Ortigão & Traub-Cseko 2003, Ramalho-Ortigão et al. 2005). Other than that, there is virtually no information regarding gene expression profiles in the midgut of this vector. Our group is investigating patterns of gene expression in L. longipalpis upon blood feeding and/or infection with Leishmania. Our approach relied initially on differential display reverse transcriptase polymerase chain reaction (DDRT-PCR) through which several cDNAs of interest were identified, including some that appear to be expressed exclusively in the midgut of L. longipalpis (Ramalho-Ortigão et al. 2001, Ramalho-Ortigão & Traub-Cseko 2003). DDRT-PCR has also been successfully used in the characterization of many mosquito tissue-specific cDNAs, including an A. gambiae midgut-specific V-ATPase (Dimopoulos et al. 1996). V-ATPase, one of the most fundamental enzymes in nature, energizes a wide variety of organelles and membranes in organisms ranging from insects to higher vertebrates (Nelson 2003). They consist of two large multi-meric domains (a membrane spanning and an intracellular), each of which composed by several subunits (Forgac 1989). Such proteins function as proton (H+) pumps, having the ability to stimulate acidification of cytoplasm in a number of cells. They promote the acidification of the midgut lumen in Bombyx mori (Wieczorek et al. 2000) and were identified and characterized in the mosquitoes Culex quinquefasciatus (Fillipova et al. 1998), A. gambiae (Dimopoulos et al. 1996), and Aedes ae-gypti (Gill et al. 1998, Zhuang et al. 1999, Weng et al. 2003). Additionally, Plasmodium gallinaceum ooki-netes were shown to selectively invade A. aegypti midgut cells in the vicinity of cells expressing high levels of V-ATPase (Cociancich et al. 1999). Later work showed that in A. stephensi, midgut invasion by P. berghei preferentially occurs through cells expressing low levels of V-ATPase (Han et al. 2000). Also, V-ATPases seem to have a role in gregarine infection of A. albopictus (Huang et al. 2006). Using DDRT-PCR we have identified a cDNA for V-ATPase subunit C from Leishmania infected L. lon-gipalpis RNA (Ramalho-Ortigão et al. 2001). In this work, we report the characterization of a full length cDNA coding for this V-ATPase subunit C of L. longipalpis isolated from a midgut expression library established with RNA isolated 6 h after a blood meal. The cDNA shows a high degree of identity with its counterparts identified in many insects including the mosquito vectors A. gambiae and A. aegypti, Drosophila melanogaster, Apis mellifera, and the lepidopterans B. mori and Man-duca sexta. This cDNA is up-regulated in the midgut of blood fed L. longipalpis, and has a modulated expression during insect development. In sand flies, acidification of the midgut following digestion of the blood meal has been incriminated as one of the mechanisms inducing metacyclogenesis of Leishmania and parasite migration towards the foregut (Gontijo et al. 1998), which are fundamental events for subsequent infection of vertebrates. A potential role for V-ATPase in this process remains to be assessed. MATERIALS AND METHODS Sand flies - Sand flies were originally captured at Santarém (Northern Brazil) and maintained in colony at the Evandro Chagas Institute, Belém. Infection with L. i. chagasi amastigotes was done with parasites harvested from hamster-infected spleen, homogenized in blood (approximately 106/ml) and fed to L. longipalpis adult females using one-day old chick skin membrane. Fully engorged females were dissected 72 and 96 h after feeding and checked for Leishmania in the midgut. Infected or non-infected insects were used for RNA extraction with TrizolTM (Life Technologies, Grand Island, NY, US) according to instructions. DDRT-PCR - RNA purification and DDRT-PCR were performed as described in Ramalho-Ortigão et al. (2001). A differentially amplified band was cloned into pCDNA3 and sequenced. This 220bp cDNA was shown to be similar to V-ATPases of different sources and subsequently used as probe for the detection a complete cDNA from an expression library. Expression libraries - Total RNA (100-200 ng) purified from midguts dissected 6 h after blood feeding were used for synthesis of a cDNA library with the lZAP kit from Stratagene (US), following manufacturer's instructions. The original library was amplified (Sambrook et al. 1989) and the supernatant containing 1010 pfu (plaque forming units)/ml used to transfect freshly grown XL1blue cells (Stra-tagene, US). Plaques were blotted onto nylon membranes, treated as described (Sambrook et al. 1989), and hybridized to the 220pb V-ATPase cDNA fragment radioactively labeled using an Amersham-Pharmacia (US) Random Priming kit according to manufacturer's protocol. Blots were washed (3 x 0.1% SSC-0.2 XSSC at 50oC) and exposed to X-ray film. Positive plaques were picked, transferred to SM buffer, and re-plated for a second round of screening. Finally, positive plaques were used for excision of the pBlu-escript plasmid and sequenced at the PDTIS/Fiocruz sequen-cing platform, in an Applied Biosystems 3730 DNA analyzer. RT-PCR - Total RNA was extracted with Trizol reagent according to manufacturer's protocol from groups of 20 fully engorged females at 2, 6, 12, 24, 48, 72, 96, and 168 h after blood feeding, or 72 h after artificial L. i. chagasi infection. Total RNA also was extracted from unfed females (0 h), males, eggs, the four larval stages, and pupae. Up to 5 mg of total RNA from each time point was used for 1st strand cDNA synthesis, performed with the First-Strand cDNA Synthesis kit (Amersham). Each cDNA was amplified during PCR reactions containing 0.8 mM of each specific primer set in a 25 ml reaction. PCR was carried out using the following conditions: 96oC/3 min, followed by 35 cycles at 96oC/30 s, 57oC/45 s, and 72oC/45 s, and a final extension of 72oC/5 min. The oligonucleotides for V-ATPase were: LlV ATPase-F 5' GTC AGC CTA CAA CAG CCT CAA 3' and LlVATPase-R 5'AGT TTA CGG GCA ATC CAT ACC 3'. As constitutive expression control histone primers were used: His-F 5' GAA AAG CAG GCA AAG ACT CC 3' and His-R 5' GAA GGA TGG GTG GAA AGA AG 3'. PCR products were separated in 2% ethidium bromide-stained agarose gels. Multiple alignment - Using BLAST search for similarity in GenBank, several V-ATPase sequences from different insects were identified and aligned with ClustalW. V-ATPase sequences used for the alignment were: B. mori (ABD36144.1); M. sexta (CAB55498.1); A. mellifera (XP_395359.3); D. melanogaster (AAB62571.1); A. aegypti (ABF18462.1); A. gambiae (XP_556516.1). The output format of ClustalW was submitted to BOX SHADE program to generate the alignment figure. Phylogenetic tree - A phylogenetic tree was obtained from the V-ATPase multiple alignment (ClustalW) from different insects (B. mori, M. sexta, A. mellifera, D. melanogaster, A. aegypti, A. gambiae) using MEGA3 software. The phylogeny test used was the Neighbor-joining that was carried out with Poisson correction and bootstrap with 10,000 replications. Southern blot - Genomic DNA was extracted from adult insects using Genomic Prep Cell and Tissue DNA Isolation Kit (Amersham-Pharmacia) according to manufacturer's instructions. DNA (5 mg) was digested with the following enzymes: BamHI, BglII, EcoRI, HindIII, PvuII, PstI, and SalI, according to manufactu-rer's instructions. DNA was separated on 0.8% agarose gel, transferred to nylon membrane and hybridized to a 220 bp V-ATPase fragment previously identified by DDRT-PCR (Ramalho-Ortigão et al. 2001) labelled with a32P-dCTP using Megaprime DNA Labelling System (Amersham Life Science, US). Hybridization was done for 1 h with QuickHyb solution (Stratagene) at 60oC, blots were washed three times, 20 min each, with 0.5% SDS-0.5XSSC (Sambrook et al. 1989) at 56oC and exposed to film (X-OMAT Kodak, US) for 48 h at - 70oC. RESULTS Cloning of the V-ATPase subunit C from L. longi-palpis - We have identified and sequenced a cDNA (Fig. 1), coding for the subunit C from the sand fly L. longipalpis V1 domain of V-ATPase, by screening a midgut expression library established with RNA purified from flies dissected 6 h after blood feeding. Analysis of this cDNA revealed a high degree of similarity with many different V-ATPase subunits C, specially the ones from A. gambiae, A. aegypti, A. mellifera, D. melanogaster, B. mori, and M. sexta (Fig. 2). Subsequent analysis of the putative protein coded by this cDNA showed biochemical characteristics similar to those of its counterparts from the afore mentioned insects: putative isoeletric point (pI) of near 8.0, an estimated deduced protein molecular weight (MW) of 40kDa and a hydrophobicity/hydrophilicity profile that indicates a cytoplasmic location for the protein (i.e. highly hydrophilic) (not shown). Multiple alignment - Multiple alignment of the amino acid sequences from different insects and from the deduced L. longipalpis sequence demonstrates a well conserved protein (Fig. 2). There are several single amino acids differences among the sequences analyzed. Interestingly, the A. gambiae sequence has a large insertion absent in the other insects, and a small polymorphic site on the C terminus of the proteins. The vacuolar ATPase subunit C domain occupied almost the entire sequence. Phylogenetic tree - The molecular evolution of V-ATPase genes demonstrates a high level of conservation and a correlation with the taxonomy of the selected species (Fig. 3). The tree shows two main groups, the first one represented by the order Lepidoptera (B. mori, M. sexta) and the second by Hymenoptera and Diptera orders. Within Diptera it is possible to distinguish another division at the suborder level, the Brachycera (D. melanogaster) being separated from the Nematocera species (L. longipalpis, A. aegypti, A. gambiae). Southern blot - Genomic DNA of L. longipalpis digested with various restriction enzymes and hybridized to a fragment of the VATPase cDNA showed a pattern of hybridization compatible with one or two genes (Fig. 5). The hybridization to the PvuII digest originates a large double band, which might be explained by the presence of two gene copies, or by a slight size difference of the gene alleles. We have previously found a size difference of chitinase alleles in L. longipalpis (unpublished results). RT-PCR - Fig. 4A shows the results of RT-PCR using cDNA prepared from RNAs extracted from L. longipalpis in different stages of development, namely eggs, larvae from the first (L1), second (L2), third (L3), and fourth (L4) instar, pupae, and male and female adults fed on sugar. There is a low level of V-ATPase expression in eggs, followed by a slight decrease of expression in L1 larvae and a gradual increase in expression up to pupae. In non-blood fed, the highest expression is found in adult males. In Fig. 4B the expression of V-ATPase was investigated in relation to blood feeding. An increase in V-ATPase expression was detected as early as 2 h post blood meal (PBM), continuing up to 24 h PBM. From the apparent peak at 24 h PBM, expression of V-ATPase was down regulated beginning at 48 h PBM and by 96 h PBM the expression reached its lowest levels. The expression levels at 72 h PBM appear to be equivalent to what was detected for L1 larvae. Additionally, no difference in V-ATPase expression was detected when we compared un-infected versus infected blood feeds, 72 h following a blood meal. DISCUSSION V-ATPases are heteromeric proteins that, in general, function as proton pumps (reviewed in Harvey & Nelson 1992). These enzymes, found in all eukaryotes, are composed of catalytic and transmembrane spanning domains, and possess a broad range of biological functions (Merzendorfer et al. 1997, Forgac 1998, Nelson 2003). In insects V-ATPases are able to perform roles such as membrane energization (Klein 1992) and acidification of cellular compartments or even whole insect's midgut (Jäger et al. 1992, Wieckzorek et al. 1996). V-ATPases cytoplasmic (F1) and membrane (F0) domains, each consists of several subunits (Harvey & Nelson 1992). The subunit C, although not required for enzymatic activity under certain conditions (Puopolo et al.1992), appears to be important for the assembly of the F1 domain (Ho et al. 1993). We have cloned a complete cDNA coding for the V-ATPase subunit C of the sand fly vector L. longipalpis. This full length cDNA clone was obtained by screening a midgut expression library with a previously identified DDRT-PCR fragment of approximately 220 bp obtained from RNA of insects artificially infected with L. braziliensis (Ramalho-Ortigão et al. 2001). The complete cDNA codes for a 386 amino acid predicted protein with an estimated molecular weight of 40 kDa. The L. longipalpis V-ATPase (LlVATPase) shows a high degree of identity and similarity, based on amino acid comparison, with their counterparts from A. aegypti and A. gambiae, D. melanogaster, M. sexta. LlVATPase possesses three putative glycosylation sites, similar to what is predicted for A. americanum and M. sexta. In contrast, Drosophila V-ATPase subunit C appears to have four possible glycosylation sites (not shown). The amino acid and phylogenetic analysis of V-ATPase subunit C from several insects demonstrated a well-conserved protein with a small polymorphic site at the C terminus. It is interesting to note that all Nematocera species analyzed in this study are blood-feeding insects with their V-ATPases displaying a considerable evolutionary distance from those of non-blood feeding insects. Such divergence of V-ATPase C from blood feeding to non-blood feeding insects may point to a link between blood feeding behavior and/or physiology and the molecular evolution of the V-ATPase C gene. Southern blot analysis indicates possibly only one copy of the V-ATPase subunit C gene in L. longipalpis similar to what was observed for M. sexta (Merzendorfer et al. 2000). In A. aegypti, although there is a single copy of the V-ATPase A subunit, the presence of more than one copy of the subunit C gene appears to be the case (Gill et al. 1998). The expression of V-ATPase appears to be up-regulated in L. longipalpis after a blood meal. In the mosquito A. gambiae, the principal vector of human malaria, a midgut-specific V-ATPase, whose expression is apparently altered by blood feeding, has been identified (Dimopoulos et al. 1996). Analysis of V-ATPase subunit C expression throughout L. longipalpis development suggests a stage-specific regulation. While low level of expression was observed in eggs, larval stages L1 through L3, a high level of expression was seen in L4 stage larvae and pupae, approximately double of that seen in L1 larvae. These results are in contrast to what was detected for A. gambiae (Dimopoulos et al. 1996), where intermediate larval stages display the highest expression of V-ATPase, and a lower expression is seen in pupae. V-ATPase expression has also been shown in pupae and fourth instar larvae of C. quinquefasciatus, and the protein was detected in fourth instar larvae of both C. quinquefasciatus and A. aegypti (Filippova et al. 1998). However in these studies no data on the profile of V-ATPase expression of other developmental stages were obtained. Additionally, the expression of V-ATPase C is up-regulated in L. longipalpis following a blood meal, with levels approximately twice as high at 6 h after blood feeding, as compared to unfed females. Interestingly, the level of expression of this gene is approximately three times higher in males than females fed on sugar indicating an important physiological role for this molecule in L. longipalpis males. In L. longipalpis, re-acidification of the midgut following digestion of a blood meal has been linked to metacyclogenesis of Leishmania within the sand fly (Gontijo et al. 1998). Before a blood meal, midgut pH was measured at approximately pH 6 and the hindgut at a pH lower than pH 6. Upon taking a blood meal, pH in the gut increases as a result of the pH 7.4 of blood. After complete digestion, pH may decrease to levels prior to the blood meal. Gontijo et al. (1998) proposed that the midgut reacidification would stimulate metacyclogenesis of Leishmania taken with an infective blood meal as seen with cultured parasites, where low pH induces metacyclogenesis (Zakai et al. 1998). A possible role for V-ATPases in events leading to metacyclogenesis of Leishmania remains to be elucidated. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07080f1.jpg] [oc07080f5.jpg] [oc07080f4.jpg] [oc07080f3.jpg] [oc07080f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}