 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
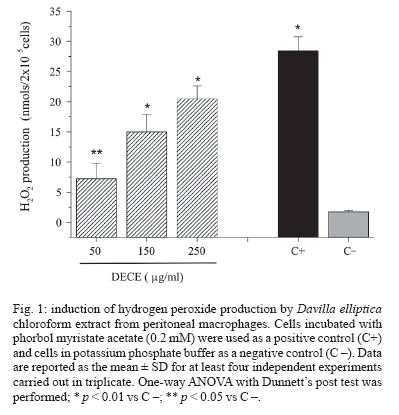
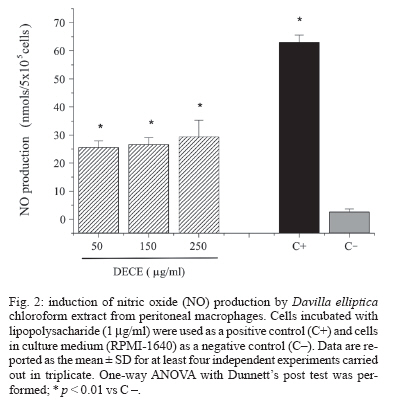
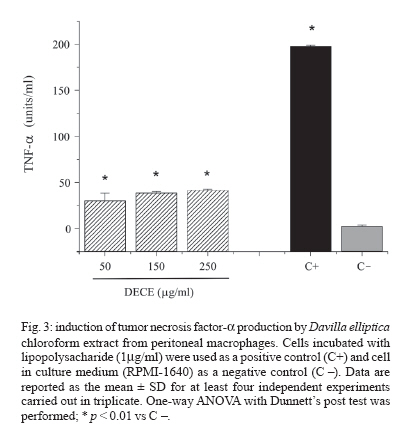
Memórias do Instituto Oswaldo Cruz, Vol. 102, No. 6, 2007, pp. 769-772 Immunological and microbiological activity of Davilla elliptica St. Hill. (Dilleniaceae) against Mycobacterium tuberculosis Flávia Cristine Mascia Lopes, Marisa Campos Polesi Placeres, Cleso Mendonça Jordão Junior*, Célio Takashi Higuchi*, Daniel Rinaldo**, Wagner Vilegas**, Clarice Queico Fujimura Leite*, Iracilda Zeppone Carlos/+ Departamento de Análises Clínicas *Departamento de Ciências Biológicas **Departamento de Química Orgânica, Unesp, R. Expedicionários do Brasil 1601, 14801-902 Araraquara, SP, Brasil Financial support: Fapesp-Biota program (02/05503-6), CNPq Received 12 February 2007 Accepted 16 July 2007 Code Number: oc07123 Mycobacterium tuberculosis is responsible for over 8 million cases of tuberculosis (TB) annually. Natural products may play important roles in the chemotherapy of TB. The immunological activity of Davilla elliptica chloroform extract (DECE) was evaluated in vitro by the determination of hydrogen peroxide (H2O2), nitric oxide (NO), and tumor necrosis factor-a (TNF-a) release in peritoneal macrophages cultures. DECE was also tested for its antimycobacterial activity against M. tuberculosis using the microplate alamar blue assay. DECE (50, 150, 250 µg/ml) stimulated the production of H2O2 (from 1,79 ± 0,23 to 7,27 ± 2,54; 15,02 ± 2,86; 20,5 ± 2,1 nmols) (means ± SD), NO (from 2,64 ± 1,02 to 25,59 ± 2,29; 26,68 ± 2,41; 29,45 ± 5,87 µmols) (means ± SD) and TNF-a (from 2,44 ± 1,46 to 30,37 ± 8,13; 38,68 ± 1,59; 41,6 ± 0,90 units/ml) (means ± SD) in a dose-dependent manner and also showed a promising antimycobacterial activity with a minimum inhibitory concentration of 62,5 µg/ml. This plant may have therapeutic potential in the immunological and microbiological control of TB. Key words: Davilla elliptica - antimycobacterial activity - immunological activity Mycobacterium tuberculosis is a serious threat to humankind, with over 8 million cases of tuberculosis (TB) annually (Raupach & Kaufmann 2001). Most healthy individuals are able to control TB infection with a vigorous immune response, halting the progression of the disease, but not necessarily eradicating the microorganism (McKinney 2000). The bacterium resides within macrophages, allowing them to resist the antimicrobial effector mechanisms of the host (Raupach & Kaufmann 2001). The major effector molecules of antibacterial defense in macrophages include reactive oxygen intermediates (ROI) and reactive nitrogen intermediates (RNI) (Nathan & Shiloh 2000). Hydrogen peroxide (H2O2), one of the ROI generated by macrophages in a reaction catalyzed by an NADPH oxidase, was the first identified effector molecule that mediated mycobacteriocidal effects of mononuclear phagocytes (Walker & Lowrie 1981). Nitric oxide (NO), formed by the action of the inducible form of nitric oxide synthase (iNOS), reacts with oxygen radical forming RNI. NO and related RNI have been reported to possess antimycobacterial activity (Kwon 1997). M. tuberculosis strongly induces the release of several cytokines during infection. Tumor necrosis factor-a (TNF-a) is a cytokine that plays multiple roles in immune and pathologic responses in tuberculosis, also required for acute infection control (Flynn et al. 1995). It plays a major role in the recruitment of inflammatory cells to the site of infection and in the formation and maintenance of granulomas (Gaemperli et al. 2006). Davilla elliptica St. Hill. (Dilleniaceae) is a plant popular known as cipó-caboclo. Although there are not many studies focused on this plant, it is commonly used as tonic, astringent, and laxative in the Brazilian folk medicine (Rodrigues & Carvalho 2001). In this study, the immunological activity of D. elliptica chloroform extract (DECE) was evaluated in vitro by the determination of H2O2, NO, and TNF-a release in peritoneal macrophages cultures. DECE was also tested for its anti-mycobacterial activity against M. tuberculosis using the microplate alamar blue assay (MABA). Plant material and samples - D. elliptica leaves were collected at Porto Nacional, Tocantins, Brazil (no. 4583 - Tocantins University Herbarium). The leaves were dried at 40ºC and then macerated with chloroform. After solvent evaporation, the extract was obtained. The extract was first solubilized in dimethyl sulfoxide (DMSO) and then diluted in an appropriated culture medium, RPMI-1640 for the immunological assays (50, 150, 250 µg/ml) and Middlebrook 7H9 for the determination of antimyco-bacterial activity (62,5-4000 µg/ml). Animals - Experiments involving Swiss mice (6-8 weeks old, 18 to 25 g) were performed in accordance with the regulations of Research Ethics Committee (# 01/2005), Faculty of Pharmaceutical Sciences, Unesp, São Paulo, Brazil. Peritoneal macrophages - Thioglycollate-elicited peritoneal exsudate cells (PEC) were harvested from Swiss mice using 5 ml of sterile PBS, pH 7.4. The cells were washed twice by centrifugation at 200 g for 5 min at 4ºC and ressuspended in an appropriate medium for each test. MTT assay was performed to assure the viability of the cells in the presence of DECE (Mosmann 1983) (data not shown). Measurement of H2O2 production - H2O2 release was measured using the horseradish peroxidase-dependent phenol red oxidation microassay according to Pick and Mizel (1981). Cells incubated with phorbol myristate acetate (PMA) 0.2 mM only were used as a positive control. Measurement of NO production - NO production induced by DECE was determined by assaying culture supernatants for nitrite using Griess reagent according to Green et al. (1982). Cells incubated with 1µg/ml of lipopolysacharide (LPS) from Escherichia coli O111:B4 were used as a positive control. Measurement of TNF-a production - The determination of TNF-a in the supernatants was based in its property to destroy L929 tumoral cell line (Carlos et al. 1994). LPS (1 µg/ml) was used as a positive control. Determination of antimycobacterial activity (MABA technique) - The minimum inhibitory concentration (MIC) of DECE was determined against M. tuberculosis H37Rv (American Type Culture Collection 27294) (107/ml) in Middlebrook 7H9 medium using the MABA (Collins & Franzblau 1997). The classic antimycobacterial drug isoniazide (INH) (Sigma) (0,015-1 µg/ml) was used as a reference drug. Statistical analysis - The results are expressed as means ± SD of five experiments. One-way ANOVA with Dunnett's post test was performed using GraphPad InStat (San Diego, California, US) with the level of significance set at p < 0.05. In recent years, emerging multiple drug resistance has become a major threat and thus calls for an urgent search for new and effective treatments for this deadly disease. Naturally occurring compounds as extracts from plants have indicated that inhibitory activity against M. tuberculosis is widespread in nature (Okunade et al. 2004). In the present study, it was demonstrated that addition of different concentrations of DECE extract (50, 150, 250 µg/ml), to cultured macrophages, triggered the production of H2O2 (from 1,79 ± 0,23 to 7,27 ± 2,54; 15,02 ± 2,86; 20,5 ± 2,1 µmols) (means ± SD) (Fig. 1), NO (from 2,64 ± 1,02 to 25,59 ± 2,29; 26,68 ± 2,41; 29,4 5,9 µmols) (means ± SD) (Fig. 2) and TNF-a (from 2,44 ± 1,46 to 30,37 ± 8,13; 38,68 ± 1,59; 41,6 ± 0,9 units/ml) (means ± SD) (Fig. 3) in a dose-dependent manner. The discrete but significant production of the three studied mediators (H2O2, NO and TNF-a) by DECE stimulated macrophages is very important, since macrophages produces several effector molecules that can enhance or restore the ability of the immune system to fight against TB infection. It is well know that H2O2 is an effective microbial disinfectant, destroying pathogens by an oxidation process that results in protein denaturation (Hughes et al. 2003). Studies have shown that M. tuberculosis infection induces the accumulation of macrophages in the lung and in the ascitic fluid and also H2O2 production in patients with pulmonary tuberculosis and tuberculous ascites (Swamy et al. 1988, Raja 2004). Phagocytes kill intracellular organisms during an initial oxidative phase dependent on NADPH oxidase, followed by a prolonged nitrosative phase, during which bacterial growth is inhibited by iNOS (Nathan & Shiloh 2000). There are several potential mechanisms that can explain how NO may affect microbial life-cycle. NO and other RNI can modify bacterial DNA, protein and lipids at both the microbial surface and intracellularly. They can alter cytokine production and induce or prevent apoptosis of host cells by controlling caspase activity (Raupach & Kaufmann 2001). NO can also interact with accessory protein targets, resulting in enzymatic inactivation or other protein malfunctions (Gow et al. 1999). Besides that, NO reacts with superoxide to form a strong oxidant, peroxynitrite (ONOO-). ONOO- mediates ni-trosylation of tyrosine residues, and therefore has the potential to disrupt tyrosine phosphorylation-dependent signaling pathways (Linares et al. 2001). Studies in mouse infection models have clearly revealed that TNF-a is a crucial component of both the antibacterially protective and the inflammatory immune response to M. tuberculosis. It is necessary for optimal co-ordination of both the differentiation of specific T cells to secrete the appropriate T helper 1 cytokines and the development of granulomas in which activated macrophages restrict mycobacterial growth (Ehlers 2003). TNF-a is required for control of latent TB and it is also a key element for activating macrophages to produce iNOS and thus in maintaining the pathway for generating NO and preventing reactivation of the disease (Adams et al. 1995). In our research, DECE exhibited a antimycobacterial activity (MIC = 62,5 µg/ml), indicating that the extract could be an important source of bactericidal compounds against M. tuberculosis. Although still being less potent than the reference drug INH (MIC = 0,03 µg/ml), we have to consider that D. elliptica extract was tested and not pure compounds as the study conducted by Akihisa et al. (2005). In this research, 15 pure compounds with MIC values of 4-64 µg/ml were isolated from the flower extract of Chrysanthemum morifolium and showed high activity against M. tuberculosis, being considered valuable from the viewpoint of anti-TB. Recent advances in understanding the host-pathogen interaction created new approaches for the development of new antimycobacterial agents. In the present work, it was demonstrated that DECE exhibited promising results suggesting that this plant may have therapeutic potential in the immunological and microbiological control of TB. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07123f2.jpg] [oc07123f1.jpg] [oc07123f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}