 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
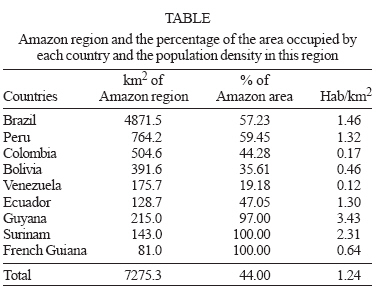
Memórias do Instituto Oswaldo Cruz, Vol. 102, No. Suppl. I, 2007, pp. 47-55 Chagas disease in the Amazon Region Hugo Marcelo Aguilar, Fernando Abad-Franch*, João Carlos Pinto Dias**, Angela Cristina Veríssimo Junqueira***, José Rodrigues Coura***/+ Organismo Andino de Salud, Quito, Ecuador *Instituto Leônidas & Maria Deane-Fiocruz, Manaus, AM, Brasil **Centro de Pesquisas René Rachou-Fiocruz, Belo Horizonte, MG, Brasil ***Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil Finalcial support: The WHO TDR Special Programme (ID # A20441), Fiocruz, ECLAT, Fapeam, CNPq (Proc. 400274/2006-0) Received 4 July 2007 Accepted 3 September 2007 Code Number: oc07134 The risk that Chagas disease becomes established as a major endemic threat in Amazonia (the world's largest tropical biome, today inhabited by over 30 million people) relates to a complex set of interacting biological and social determinants. These include intense immigration from endemic areas (possibly introducing parasites and vectors), extensive landscape transformation with uncontrolled deforestation, and the great diversity of wild Trypanosoma cruzi reservoir hosts and vectors (25 species in nine genera), which maintain intense sylvatic transmission cycles. Invasion of houses by adventitious vectors (with infection rates > 60%) is common, and focal adaptation of native triatomines to artificial structures has been reported. Both acute (~ 500) and chronic cases of autochthonous human Chagas disease have been documented beyond doubt in the region. Continuous, low-intensity transmission seems to occur throughout the Amazon, and generates a hypoendemic pattern with seropositivity rates of ~ 1-3%. Discrete foci also exist in which transmission is more intense (e.g., in localized outbreaks probably linked to oral transmission) and prevalence rates higher. Early detection-treatment of acute cases is crucial for avoiding further dispersion of endemic transmission of Chagas disease in Amazonia, and will require the involvement of malaria control and primary health care systems. Comprehensive eco-epidemiological research, including prevalence surveys or the characterization of transmission dynamics in different ecological settings, is still needed. The International Initiative for Chagas Disesae Surveillance and Prevention in the Amazon provides the framework for building up the political and scientific cooperation networks required to confront the challenge of preventing Chagas disease in Amazonia. Key words: Chagas disease - Trypanosoma cruzi - epidemiology - surveillance - Amazonia AMAZONIA: OVERVIEW OF A COMPLEX BIOME The greater Amazon lies on the largest hydrographical basin of the world, comprising 44% of the South American subcontinent (Fig.1, Table). It contains about 20% of all liquid freshwater on the planet, distributed over 300,000 km2 of aquatic ecosystems. The Amazon River runs for ~ 6270 km, receiving waters from ~ 10,000 highly heterogeneous tributaries. The humid tropical climate of Amazonia varies significantly in space and time, with mean annual rainfall ranging from 1200-2000 mm in the northern and southern fringes of the region to up to 6000 mm in the vicinity of the Andes and in the Brazilian states of Pará and Amapá. The vast Amazonian fluvial network irrigates the largest tropical biome on Earth (6.5 million km2). Far from being homogeneous, the Amazon biome takes the shape of a complex mosaic of highly diverse ecosystems, including over 25 terrestrial eco-regions comprising no less than 180 distinct land systems. Tropical humid forests are widely predominant, but certain semi-arid formations, sandy soil campinas and campinaranas, várzea pioneer formations, and a variety of anthropic landscapes are also present. The forests and savannahs of the Amazon harbour the largest biological diversity (in absolute terms) on the planet (cf. Abad-Franch et al. 2006). Over 30 million people live in the Amazon region (< 5 inhabitants/km2). In the Brazilian Amazon alone, over 6 million people live in traditional, mostly rural societies. An estimated 260,000 Amerindians, speaking some 170 native languages, live in the region. Caboclos, ribeirinhos, rubber-tapers, and other traditional colonists typically establish themselves in small communities along river banks, depending on subsistence farming, fishing, and forest extractivism. During the second half of the XX century, government policies of colonization and development, linked to huge infrastructure projects, fostered unprecedented immigration movements towards the Amazon. In Brazil, population density increased from 1.5 to 4.5 inhabitants/km2 between 1950 and 2000. Road opening, especially in the southern fringe of the region, set off a process of disordered colonization that resulted in annual deforestation rates between 15,000 (1993) and almost 25,000 km2 (2002). By 1970, about 65% of the population of the Brazilian Legal Amazon lived in rural settings; 30 years later, 70% of the 21 million inhabitants of the region lived in cities (cf. Abad-Franch et al. 2006). The forests with their biological diversity, the stocks of strategic resources (minerals, water, energy, genes, chemicals, timber, and land), and native peoples and cultures threatened by a homogenizing civilization are among the major concerns underlying the current global geopolitical interest on Amazonia. It is in this context that Amazonian nations have changed their view of the region over the last two decades. Differential investment and tax incentive policies in several countries or international collaboration initiatives in various domains (politics, security, economics and trade, energy, science and technology, higher education, etc.) are all the expression of this trend. The new international dimension of the region has effectively relocated Amazonia from the periphery of each individual country (with Amazonian territories typically less developed than the rest) to the center of the continent. However, and despite substantial efforts of some countries over recent years, social policy planning in the region is still far from ideal. International health initiatives such as those aimed at reducing the burden of endemic infectious diseases may in fact have a wider impact than that related to its specific objectives, promoting the integration and coordination of national science and technology policies across Amazonian countries. In addition to malaria, which is highly prevalent, major communicable diseases that are common in the region include different viral infections (such as viral hepatitis and dengue fever), leishmaniasis, leprosy, and tuberculosis. The epidemiological status of human Chagas disease in Amazonia is only superficially understood. The disease used to be considered rare until it was labelled as emergent in two recent publications (Teixeira et al. 2001, Coura et al. 2002). With the advent of the International Initiative for Chagas Disesae Surveillance and Prevention in the Amazon (AMCHA), officially launched under the auspices of the PAHO-WHO in 2004, public health researchers and officials became engaged in renewed efforts to elucidate the actual significance and transmission dynamics of the disease in the Amazon region (Guhl & Schofield 2004, Anonymous 2005, PAHO 2005). In this paper we present a summary of the epidemiological status and trends of Chagas disease in Amazonia, reflecting on the relative roles of social and biological factors in the complex dynamics of Trypanosoma cruzi transmission to people across the region, and emphasize the current progress of international control-surveillance initiatives. CHAGAS DISEASE IN AMAZONIA: GENERAL PICTURE The risk that Chagas disease becomes established as a major endemic threat to Amazonian populations relates to a complex set of interacting biological and social determinants. Among the latter, intense human migration from endemic areas into the Amazon (with the possibility of unintentionally introducing foreign parasite strains and vectors) and extensive landscape transformation with uncontrolled deforestation are probably the most important. Biological factors are linked to the great diversity of wild T. cruzi reservoir hosts (marsupials, bats, rodents, edentates, carnivores, primates, etc.) and vectors (over 25 species in nine genera), which results in intense sylvatic transmission cycles (cf. Abad-Franch et al. 2006). Invasion of houses by adventitious vectors (with infection rates usually above 60%) is common, and focal adaptation of native triatomines to human dwellings has been reported from several areas. Fig.2 illustrates a widespread risk situation: a rural house in the Brazilian Amazon was built close to palm trees occupied by triatomines and marsupials both frequently infected with T. cruzi. Hundreds of acute cases of human Chagas disease have been reported from across the Amazon. Chronic forms of the disease are however considered to be relatively rare in the region. The first acute cases were reported by Floch and Tasqué (1941) and Floch and Camain (1948) from French Guiana. Shaw et al. (1969) described four further acute cases in Belém, capital city of the state of Pará, in Northern Brazil. Since then, about five hundred acute cases have been recorded, most of them in the Brazilian states of Pará, Amapá, and Amazonas (e.g., Valente & Valente 1993, Valente et al. 1999, Pinto et al. 2004, Perez-Gutierrez et al. 2006, PAHO 2006a). Overall prevalence of infection is relatively low (1-3%), but seems to be substantially higher in some subregions (Camargo et al. 1984, Dias & Coura 1997). Serological studies carried out by the Fundação Nacional de Saúde (Funasa) from 1975 to 1980 and by Coura and his collaborators from 1971 to 2002 in the Brazilian Amazon revealed prevalence rates from ~ 2 to 13% (Camargo et al. 1984, Coura et al. 1999, 2002). Amazonian T. cruzi strains generally belong to group 1 (Z1), zymodeme Z3, and hybrid Z1/Z3 forms (Fernandes et al. 2001, Coura et al. 2002). As a rule, morbidity and mortality rates are low in Amazonia, and certainly significantly lower than in endemic regions like the Southern Cone. Still, isolated, well-documented records of chronic cardiopathy, heart enlargement, and death caused by Chagas disease in autochthonous patients exist in the region (Albajar et al. 2003, Xavier et al. 2006). The identification and reporting of acute cases (both isolated and related to focal outbreaks) is more common in areas with a tradition of research activities (such as Pará, Brazil). Epidemiological evidence strongly suggests oral T. cruzi transmission in many of the outbreaks of acute cases recorded in Brazil (Valente & Valente 1993, Valente et al. 1999, Pinto et al. 2004, Coura 2006, Perez-Gutierrez et al. 2006, PAHO 2006a). This epidemiological picture (relatively low prevalence, incidence, morbidity, and mortality rates but by no means absence of the disease) is also supported by data on prevalence of T. cruzi infection among blood donors and by the rarity of hospital admissions and death due to Chagas disease in the Brazilian Amazon (Dias et al. 2002). COUNTRY SUMMARIES In this section, we summarize data on the status of Chagas disease in the nine Amazonian countries provided by official technical representatives (Brazil, Colombia, Ecuador, and Peru) or ECLAT experts (Bolivia, French Guiana, Guyana, Suriname, and Venezuela). This overview is based on reports presented at the 2002 and 2005 AMCHA meetings (Dias et al. 2002, Guhl & Schofield 2004, Abad-Franch et al. 2006), with contributions from further sources and updated information. Systematized, in-depth information on the epidemiology of Chagas disease in the Amazon is clearly insufficient; well-defined research priorities and organized control programs are also lacking in all the countries of the region. Bolivia Two-thirds of the country are at risk of vector-borne Chagas disease transmission, basically because of high rates of household infestation by Triatoma infestans. Some estimates suggest that incidence reached 180,000 new cases/year in the late 1990s. At that time, at least 30% of blood donations were infected, and only a few blood banks were screening them before transfusion. Congenital transmission of the disease was also commonplace. The Amazon region (Departments of La Paz, Beni, and Pando) is considered non-endemic, even if several sylvatic vectors are present (Rhodnius pictipes, R. robustus, R. stali, Panstrongylus geniculatus, Microtriatoma trinidadensis, and Eratyrus mucronatus, among others) and immigration from endemic areas is growing. This situation clearly calls for systematic epidemiological research, including an assessment of the risk of passive introduction of T. infestans. Bolivia prioritized Chagas disease control in 1999, and set up an ambitious program for which US$ 25 million were borrowed from the Interamerican Development Bank. The aims were to spray 700,000 dwellings, to screen all blood donations, and to provide specific treatment for all acute cases and infected children under 15 years. Preliminary data indicated that by 2002 about 350,000 dwellings had been sprayed, with a 90% reduction of household infestation by T. infestans in intervention areas. After completing full coverage of the endemic area, the Bolivian program foresees the expansion of research-surveillance activities to the Amazon (Dias et al. 2002). Of particular concern are the rising trends of immigration from the endemic highlands into the Amazon, fostered by the decline of the Andean mining industry and the parallel intensification of fossil fuel extraction and cattle ranching. Focal domestic-peridomestic infestation by populations of R. stali in several sub-regions also deserves careful investigation (cf. Abad-Franch et al. 2006). Brazil While records of triatomines and infected wild reservoirs are common ever since the times of Carlos Chagas, human disease cases still seem to be relatively rare in the Brazilian Amazon. Various Rhodnius and Panstrongylus species (mainly R. pictipes, R. robustus, and P. geniculatus, often with high rates of natural infection by T. cruzi) are present in a wide variety of natural ecotopes throughout the Brazilian Amazon. Invasion of houses by adult specimens is frequently recorded, while truly domestic vector populations occur only in geographically restricted foci e.g., T. maculata in Roraima. Symptomatic disease cases are rare, but small family or neighborhood outbreaks of acute Chagas disease, probably resulting from oral transmission, have been detected in the states of Pará, Amapá, Amazonas, Maranhão, and Acre over recent years (cf. PAHO 2006a). The national serological survey (1975-1980) revealed an overall prevalence rates of 2.4% in Acre, 1.9% in Amazonas, 0.5% in Pará, 0.4% in Rondônia, 0.3% in Roraima, and 0% in Amapá, but age classes and migration history were not recorded (Camargo et al. 1984). Seroprevalence data and entomological records suggested a disproportionately high risk of Chagas disease transmission in some municipalities, justifying their compulsory inclusion in the second national prevalence survey (Valente et al. 1998, Dias et al. 2002). This new survey, now in progress, was planned to cover children less than 5 years old in 132 municipalities of the Brazilian Legal Amazon, plus over 190 municipalities in the neighboring states of Tocantins, Maranhão, and Mato Grosso. Most of the ~ 440 acute cases recorded in the Brazilian Amazon did not present clinical evidence of the portal of entry of the parasite, and tended to respond satisfactorily to specific treatment with benznidazole. Many patients lived in urban and periurban areas, and T. cruzi infection was often detected in malaria thin smears. Infection rates among blood donors are steadily decreasing, and the Ministry of Health reports that serological screening covers practically all blood donations in the region. Sporadic cases of heart enlargement and chronic cardiopathy due to Chagas disease have been recorded, but the rates of hospitalization and mortality due to the disease are relatively low. T. cruzi isolates from autochthonous cases, vectors, and natural reservoirs from Amazonia generally belong to group 1, which is more difficult to isolate and maintain in laboratory cultures. There is a tendency for conventional serological tests to yield false-positive results in the region, and confirmation of suspect cases by means of complementary techniques such as Western blot is recommended. There is no regular program for Chagas disease control in the Brazilian Amazon. The Funasa (Ministry of Health) traditionally investigated suspect cases, sporadically detecting acute T. cruzi infections in febrile patients during routine malaria diagnosis procedures throughout the Amazon region. In the mid 1990s, 3500 Funasa laboratory technicians were working in the Brazilian Amazon; they were carrying out about two million thick and thin smear examinations on febrile patients every year. As a result of decentralization policies, most of these technicians currently work for municipal health departments. The Secretaria de Vigilância em Saúde (Department for Health Surveillance) of the Ministry of Health is currently the only federal public health agency widely present throughout the Amazon. Its activities are focused on malaria and dengue control, but it has recently supported the training of malaria technicians for T. cruzi identification in blood smears. These activities involved reference centers and public health laboratories in all the state capitals of the Brazilian Amazon, and were carried out in collaboration with Médecins sans Frontières and the Department of Tropical Medicine of the Instituto Oswaldo Cruz-Fiocruz. Research institutions such as the Instituto Evandro Chagas (IEC, Ministry of Health), Instituto Nacional de Pesquisas da Amazônia (Inpa, Ministry of Science and Technology), and Fiocruz (Ministry of Health) have had leading roles in raising public awareness in relation to Chagas disease in the Brazilian Amazon. Researchers at the IEC in particular pioneered training of malaria laboratory technicians for T. cruzi diagnosis, showing that the approach was viable and increased acute Chagas disease detection rates. The effective involvement of state and municipality health agencies has however become essential in the current context of political decentralization. In particular, the implementation of sustainable control-surveillance interventions in the Amazon crucially depends on the strengthening of public health management capabilities at the regional and local levels. Colombia The Colombian Amazon region encompasses almost 40% of the country. It is mainly comprised of humid broadleaf rainforests, sparsely populated (1625 million inhabitants), very poor, and considered to be at low risk of Chagas disease. It is in the preparatory phase for epidemiological studies and control interventions (Guhl 2005). Human settlements are mainly located along the rivers, and there is a reasonable primary health network with some regional hospitals. Triatomines recorded in the region include R. robustus, R. pictipes, R. prolixus, R. brethesi, P. geniculatus, and T. maculata. Data on the clinical epidemiology of the disease are fragmentary; a few autochthonous acute cases have been recorded recently, but information on chronic disease is absent. Partial prevalence data suggest that infection rates among native populations may reach about 2% in the department of Guainía (cf. Guhl & Schofield 2004). The few regional blood banks screen donations since 1995. T. cruzi strains isolated from vectors and reservoirs belong to T. cruzi 1 and possibly to Z3; group 2 has never been recorded. T. rangeli infection has been detected in sylvatic vectors. Ethno-anthropological evidence suggests that some indigenous peoples have a long history of contact with Chagas disease vectors, which appear in their oral tradition as evil entities that live in palm trees, suck blood, and kill their human victims (cf. Guhl & Schofield 2004). The Instituto Nacional de Salud (INS) is in charge of epidemiological intelligence and public health planning in Colombia. The INS has implemented training activities for malaria microscopists and is fostering the use of regional medical services and febrile disease sentinel units for acute Chagas disease case detection and epidemiological surveillance (cf. Guhl & Schofield 2004). Ecuador The Amazon region corresponds to the eastern most part of Ecuador, and represents about one third of the country area. Tropical broadleaf rainforests predominate, but montane life zones are also present on the Andes foothills. The extremely high biological diversity of the area is also reflected in its triatomine fauna. Sylvatic populations of at least 10 species (R. pictipes, R. robustus, P. geniculatus, P. lignarius/herreri, T. dispar, T. carrioni, T. venosa, C. pilosa and E. mucronatus) have been reported from the region. Isolated records of T. dimidiata and R. ecuadoriensis in the Amazon are probably due to mislabelling of specimens collected elsewhere. Research conducted over the past 15 years in the Ecuadorian Amazon illustrates particularly well the dynamics of T. cruzi transmission to humans in the region. Following the early description of autochthonous acute disease cases from the Napo River in the late 1980s, a serological survey conducted in the northern Ecuadorian Amazon revealed that sera from about 6% of over 1000 riverine peasants, many of them native Quechuas, were reactive for anti-T. cruzi antibodies (cf. Aguilar et al. 1999). Later on, MJ Grijalva, the late J Racines and their collaborators conducted a population-based, cross-sectional survey involving 6886 individuals from 162 communities (Grijalva et al. 2003). The distribution of seroreactivity (2.4% overall) by age class and residence history showed that (i) T. cruzi transmission to humans was taking place in several communities across the region, with nearly random geographical distribution, (ii) transmission was active at the time of the survey, and (iii) transmission had most likely been constant (and infection endemic) for at least the previous 50 years. Individual risk factors included life-long residence in the Amazon, inhabiting open households, and reporting previous contact with triatomines (Grijalva et al. 2003). When the residents of one community actively participated in vector surveillance, adult triatomines were collected inside 60% of houses within a two-month period (MJ Grijalva, pers. commun.). In the northern Ecuadorian Amazon, palm tree infestation (50% out of 110 palms surveyed) by Rhodnius spp. was significantly more frequent (and the density of vector colonies significantly higher) in deforested rural areas than in preserved forests or urban settings (FS Palomeque & FA-F, unpublished data). Taken together, these data and observations are fully compatible with low-intensity, constant T. cruzi transmission to people, most likely maintained by adult triatomines that invade rural and periurban houses from nearby infested palms. These dynamics generated an epidemiological profile typical of a hypoendemic area. French Guiana Isolated detection of acute cases and serologically reactive individuals, together with the presence of more than 40,000 Brazilian workers in the country, resulted in increasing public interest on the disease. Prevalence of T. cruzi infection may reach over 5% in some areas, and a few chronic cases have been documented among Brazilian immigrants and native afro-descendants. Acute infections have also been reported in domestic dogs in Cayenne, the capital city, suggesting urban transmission. Sylvatic cycles involve many mammals (rodents, edentates, primates, opossums and others) and a rich tri-atomine fauna. At least 12 species of Rhodnius, Triatoma, Panstrongylus, Microtriatoma, and Cavernicola occur in the country, with overall natural T. cruzi infection rates over 60% (cf. Guhl & Schofield 2004). Guyana Information on Chagas disease is scarce and fragmentary. Triatomines recorded in the country include R. prolixus, R. pictipes, R. robustus, E. mucronatus, P. geniculatus, P. lignarius, and T. maculata (Carcavallo et al. 1997, Guhl & Schofield 2004). The first acute case (an Amerindian child) was recorded in 1984. Anti-T. cruzi antibodies were detected in sera from 1% of 395 Amerindian individuals, while 500 blood donations all tested negative (cf. Guhl & Schofield 2004). Peru The Peruvian Amazon, where isolated cases of Chagas disease have been recorded, is a very sparsely populated area with both lowland and montane life zones, including drier inter-Andean valleys. Chagas disease control activities have only been enforced in the southwestern departments of Tacna, Arequipa, Moquegua, and Ica, where T. infestans maintains domestic transmission cycles. Sporadic records indicate however that chronic Chagas disease is also present in the T. infestans-free areas of Northwestern Peru and the Amazon region, where no control interventions have officially been implemented (cf. Guhl & Schofield 2004). Several triatomine species occur in these areas, and at least four of them have been reported to infest human households. P. herreri-lignarius is a domestic pest in the Marañón River Valley, and R. ecuadoriensis, P. chinai, and T. carrioni colonize houses and peridomestic structures in the Andean and coastal regions. The presence of T. dimidiata, once common in houses of Northwestern Peru, has not been recorded during the last 10 years. Triatomine records from Amazonian departments include, among others, R. pictipes, R. robustus, P. geniculatus, Belminus peruvianus, and E. mucronatus (cf. Guhl & Schofield 2004). Surinam In 1993, a serological survey among 200 Amerindian people detected two infected individuals (1%), but no infected donors were identified among 500 samples. In 2001, one acute case was recorded in a residential area of Paramaribo. Triatomines reported from Suriname include M. trinidadensis, R. pictipes, R. robustus, E. mucronatus, P. geniculatus, P. rufotuberculatus, and P. lignarius (cf. Guhl & Schofield 2004). Venezuela Venezuela has a long and dynamic history of Chagas disease research and control activities (cf. Feliciangeli et al. 2003). The main endemic areas correspond to the open lands of the Llanos and the Andes foothills within the Orinoco River basin. R. prolixus is abundant in those areas, heavily infesting rural houses with palm fiber roofs. The endemic area encompassed almost 600 municipalities, with prevalence rates from 5% to over 40%, before the implementation of the national control program. The Amazon region is the least populated part of the country, and Chagas disease prevalence rates are relatively low. T. maculata is domestic and may transmit T. cruzi in some areas, and several Rhodnius species (R. pictipes, R. brethesi, and forms of R. robustus, with occasional findings of R. prolixus) are also found in the region. Other sylvatic vectors include Psammolestes arthuri, C. pilosa, E. mucronatus, P. lignarius, and P. geniculatus. Starting in the late 1950s, an exemplary house construction and improvement program was implemented in rural Venezuela. Some 500,000 households were built or improved. In parallel, extensive chemical control interventions against domestic triatomines were implemented throughout the country, concentrating on highly endemic areas. Screening of blood donations also came into general practice, reaching nearly full coverage. This control program worked continuously until the late 1980s and had a large epidemiological impact, with an overall prevalence rate of only 3% by 1985. Prevalence among children under 10 years of age fell from 20% in the early 1960s to 0.8% in the late 1990s. Regrettably, political and economic changes in the country, together with a complex decentralization process, have significantly decreased the coverage of vector control interventions. Over the last years, domestic foci of R. prolixus have reappeared, along with several acute cases of Chagas disease. Seroprevalence trends suggest that T. cruzi transmission slightly increased in the late 1900s. Sylvatic, palm tree-living R. prolixus populations are probably involved in the infestation and re-infestation of human residences in different parts of the Orinoco basin, likely contributing to residual disease transmission (cf. Feliciangeli et al. 2003). The southern distribution limits of these R. prolixus populations probably lie within the greater Amazon biome, and the possibility that they establish domestic transmission foci in the region must be carefully monitored. A form of R. robustus genetically very close to R. prolixus is occasionally collected inside houses in both the Amazon and Orinoco basins, but seems unable to thrive in artificial environments. In the absence of detailed epidemiological data, most specialists argue that the apparent absence of endemic Chagas disease in the Venezuelan Amazon needs to be thoroughly investigated. The expertise, area coverage and infrastructure of the Venezuelan malaria control program could be crucial for establishing a surveillance network for Chagas disease in the region in which the decentralized primary health care system should also play a major role. INTERNATIONAL INITIATIVES FOR THE PREVENTION OF CHAGAS DISEASE IN AMAZONIA In July 2002, an international meeting on Chagas disease surveillance in the Amazon was held in Palmari (Amazonas, Brazil) under the auspices of the ECLAT (European Community-Latin American Triatominae) network. Since then, three further meetings were held with the aim of fostering research and public health action on Chagas disease in Amazonia.The AMCHA was officially launched in Manaus, Brazil, in September 2004 (Anonymous 2005, PAHO 2005). A second AMCHA meeting was held in Cayenne, French Guiana, in November 2005 (Abad-Franch et al. 2006), and a third meeting took place in Quito, Ecuador, in September 2006 (PAHO 2006b). Delegates from the nine Amazonian countries agreed that the PAHO in Brazil should take on the tasks of the Technical AMCHA Secretariat. The initial AMCHA recommendations called for (i) an assessment of the risk of Chagas disease becoming endemic in the Amazon region, (ii) research aimed at developing surveillance and prevention interventions, and (iii) an international cooperation system for the surveillance and prevention of Chagas disease in the Amazon region (PAHO 2005). In 2005, AMCHA delegates explicitly treated Chagas disease as a public health problem in Amazonia, stressing the endemic status of T. cruzi transmission to humans in the region and defining the control of both autochthonous and imported transmission as the shared objective of national initiatives (Abad-Franch et al. 2006). Progress reported at the 2006 meeting included the implementation of epidemiological-entomological research and surveillance activities in Ecuador, Brazil, and Colombia, the training of malaria control personnel for T. cruzi diagnosis throughout the Brazilian Amazon, and the identification of strategic opportunities to improve the availability of diagnostic tests and drugs for specific treatment in regional health centers (PAHO 2006b). A new joint meeting of the Amazonian and Andean Initiatives was scheduled for the second semester of 2007 in Lima, Peru. THE EPIDEMIOLOGICAL PATTERNS OF CHAGAS DISEASE IN AMAZONIA Almost a century after Carlos Chagas described the presence of T. cruzi in the Amazon, the epidemiology of human Chagas disease in the region remains obscure. Apparently low prevalence and morbimortality rates, together with the idea that T. cruzi transmission to humans requires domestic vectors, generated the implicit consensus that the disesae did not represent a public health problem in the region (PAHO 2005). Epidemiological research was therefore slow and fragmentary, and the results, few and fragmentary too, were often taken to confirm the thesis that Chagas disease was no priority. It is evident, for instance, that the clumped spatial distribution of cases documented in the Brazilian Amazon strongly correlates with the presence of researchers with an interest on the disease, and therefore does not reflect the true epidemiological patterns. Records of only isolated cases of human Chagas disease simply reflect the fact that only isolated researchers are looking for those cases, and we cannot infer that blank areas in epidemiological maps are really free of transmission (Coura et al. 2002). Natural cycles of T. cruzi transmission are abundant and complex in the Amazon. Some 25 triatomine species (essentially sylvatic) have been reported from the region. All of them should be considered as potential vectors, but only a few (R. prolixus, R. pictipes, R. robustus, R. stali, P. geniculatus, P. herreri, and T. maculata) seem to pose a risk for the population. Under some circumstances, Amazonian sylvatic triatomines can invade houses, contaminate foodstuffs, or attack forest workers. Some populations of T. maculata, P. geniculatus, P. herreri, and R. stali are adapted to artificial ecotopes in a few areas. Prevalence of human T. cruzi infection may be estimated as 1-2%, but reaches about 5% in some places (cf. Guhl & Schofield 2004). The identification of ~ 300 acute cases associated with family outbreaks (probably inked to the ingestion of contaminated food) has changed the traditional view of the Amazon as a region free of human Chagas disease. The number of acute cases without any relation with those outbreaks (over 125) suggests that vector-borne transmission via direct contact between humans and sylvatic triatomines is more important than compulsory case reporting indicates. Autochthonous cases of dilated myocardiopathy with fatal evolution have been documented, and severe acute cases are reported with increasing frequency. Most of these cases are associated with infection by T. cruzi 1, which circulates widely both in the Amazon and in other endemic regions (Mexico, Central America, Venezuela, and Colombia) where chronic symptomatic disease is present. Human Chagas disease may be regarded in the Amazon as an aggregate of two major epidemiological profiles that overlap in space and time. T. cruzi transmission to humans depends in both cases on the capacity of active dispersal of sylvatic vectors that fly from their natural ecotopes into houses and establish contact with humans or contaminate their food. A third risk profile is related to the synanthropic trends of some vector populations in geographically restricted areas within Amazonia, and includes the possibility of introduction of domestic vector populations from outside the region. 1. In the first situation, continuous, low-intensity transmission generates a hypoendemic pattern with seropositivity rates of about 1 to 3%. This profile is widely distributed and moderately heterogeneous, and concerns most of the rural (and a fraction of the urban) population of the Amazon. Thus, between 10,000 and 30,000 people could be infected by T. cruzi in the region. The main mechanism underlying this background transmission is the sporadic invasion of domiciles by adult sylvatic vectors (mainly R. pictipes, R. robustus, and P. geniculatus), which may be promoted by the presence of palm trees near households. Deforestation, the reduction of wild vertebrate populations, the proliferation of opportunistic mammals (rodents and opossums) in degraded environments, and the introduction of electric light are other factors potentially involved. Low visibility is inherent to some of the characteristics of these dynamics (low political weight of the populations involved, inadequate health resources, low absolute incidence, absence of house colonization by vectors, etc.). Thus, most cases probably go undiagnosed (and untreated), and information on clinical patterns and evolution is scarce. 2. The second profile involves focal, relatively intense transmission in discrete subregions. Most reported cases are linked to piaçava fiber extractivism or to the consumption of contaminated fruit juices. The fibers of the palm tree Leopoldinia piassaba are used in the manufacture of brooms and handicrafts. Fiber collection is a relevant economic activity in the middle-upper Negro River (Brazil, Colombia, and Venezuela). Whole families travel for days in small boats to reach the huge natural palm tree stands (piaçavais) that grow deep in the forests of northwestern Amazonia. They live there for several months, sleeping in open-walled, thatched huts surrounded by palms infested by R. brethesi, a particularly aggressive triatomine. Fibre collectors and their families are often attacked by these vectors, and the prevalence of anti-T. cruzi antibodies in this group reaches 5% (Coura et al. 2002). The massive consumption of agro-extractivist forest products without sanitary certification is probably involved in the family outbreaks of acute Chagas disease reported from different areas of central and eastern Amazonia. Fruit juices apparently become contaminated when infected triatomines are ground in the fruit-processing machines. This probably results in oral transmission, with high infective loads leading to severe acute disease (cf. PAHO 2006a). 3. Several foci of domestic vector populations have been described in different sub-regions throughout the Amazon: T. maculata in both rural and urban areas of Roraima, Brazil, P. geniculatus in Pará, Brazil, R. stali in the Alto Beni, Bolivia, and P. herreri in the Marañón Valley, Peru. The epidemiological risk associated with these situations has not been properly assessed. The risk of introduction of domestic vectors is also a concern in several areas. The main candidate species are R. prolixus, T. infestans , and T. dimidiata. The presence of R. prolixus in the Amazon-Orinoco transitional region is particularly worrying. This efficient vector is adapted to houses in different ecological zones from Mexico to Colombia and Venezuela, and may spread southwards following land transport routes linking Venezuela with Brazil and the Guyanas. T. infestans reached the southern limits of Amazonia in Brazil, but was subsequently eliminated from the area. Sylvatic populations occur in Bolivian Andean valleys that drain into the Amazon, but both empirical data and ecological models suggest that this species has a limited ability to survive in the humid tropics (Gorla 2002). T. dimidiata is an ecologically versatile species that can adapt to very diverse climates and ecotopes. Domestic populations occur west of the Andes in Colombia, Ecuador, and some parts of Venezuela, and could easily reach Amazonia following road networks. CONCLUSIONS A remarkable diversity of T. cruzi vectors and reservoirs occur in the Amazon. They are widely distributed geographically, occupy many different habitats and ecotopes, and are frequently infected by the parasite. The occurrence of both acute and chronic cases of autochthonous human Chagas disease has been documented beyond any doubt in the region. Continuous, low-intensity transmission seems to occur throughout the Amazon, but discrete foci exist in which transmission is more intense and prevalence rates higher. The emergence and expansion of endemic Chagas disease in Amazonia is linked to the complex ecological and social dynamics that interact in the process of occupation of this huge forested region, today inhabited by over 30 million people. Avoiding further dispersion and intensification of endemic transmission of Chagas disease in the Amazon requires the establishment of a preventive surveillance system capable of diagnosing and treating autochthonous and imported cases. The effective involvement of malaria control services and the primary health care system will be essential for achieving this goal. The need for comprehensive eco-epidemiological research, including prevalence surveys, the characterization of transmission dynamics in different ecological settings or the monitoring of synanthropic behavioral trends among autochthonous vectors, must also be emphasized. The international AMCHA initiative provides the framework for building up the political, scientific, technological, and economic cooperation networks required to confront the challenge of preventing Chagas disease in Amazonia. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07134t1.jpg] [oc07134f2.jpg] [oc07134f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}