 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No. Suppl. I, 2007, pp. 113-122 Chagas disease: what is known and what is needed A background article José Rodrigues Coura Laboratório de Doenças Parasitárias, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, Brasil Financial support: CNPq (Proc. 400274/2006-0) Received 3 August 2007 Accepted 3 September 2007 Code Number: oc07142 Chagas disease began millions of years ago as an enzootic disease of wild animals and started to be transmitted to man accidentally in the form of an anthropozoonosis when man invaded wild ecotopes. Endemic Chagas disease became established as a zoonosis over the last 200-300 years through forest clearance for agriculture and livestock rearing and adaptation of triatomines to domestic environments and to man and domestic animals as a food source. It is estimated that 15 to 16 million people are infected with Trypanosoma cruzi in Latin America and 75 to 90 million people are exposed to infection. When T. cruzi is transmitted to man through the feces of triatomines, at bite sites or in mucosa, through blood transfusion or orally through contaminated food, it invades the bloodstream and lymphatic system and becomes established in the muscle and cardiac tissue, the digestive system and phagocytic cells. This causes inflammatory lesions and immune responses, particularly mediated by CD4+, CD8+, interleukin-2 (IL) and IL-4, with cell and neuron destruction and fibrosis, and leads to blockage of the cardiac conduction system, arrhythmia, cardiac insufficiency, aperistalsis, and dilatation of hollow viscera, particularly the esophagus and colon. T. cruzi may also be transmitted from mother to child across the placenta and through the birth canal, thus causing abortion, prematurity, and organic lesions in the fetus. In immunosuppressed individuals, T. cruzi infection may become reactivated such that it spreads as a severe disease causing diffuse myocarditis and lesions of the central nervous system. Chagas disease is characterized by an acute phase with or without symptoms, and with entry point signs (inoculation chagoma or Romaña's sign), fever, adenomegaly, hepatosplenomegaly, and evident parasitemia, and an indeterminate chronic phase (asymptomatic, with normal results from electrocardiogram and x-ray of the heart, esophagus, and colon) or with a cardiac, digestive or cardiac-digestive form. There is great regional variation in the morbidity due to Chagas disease, and severe cardiac or digestive forms may occur in 10 to 50% of the cases, or the indeterminate form in the other asymptomatic cases, but with positive serology. Several acute cases have been reported from Amazon region most of them by T. cruzi I, Z3, and a hybrid ZI/Z3. We conclude this article presenting the ten top Chagas disease needs for the near future. Key words: Chagas disease - triatomines - Trypanosoma cruzi - pathogenesis - cinical forms - morbidity-mortality Natural history, origin, and distribution of Chagas disease The natural history of Chagas disease began millions of years ago as an enzootic disease of wild animals, which still persists in enzootic areas like the Amazon region. When man ventured into the natural ecotopes of the infection, it started to be transmitted accidentally to man as an anthropozoonosis. As a result of forest clearance for extensive agriculture and livestock rearing in Latin America over the last 200 to 300 years, triatomines that were unable to feed because of the displacement of wild animals started to colonize areas around and within human homes. They have become adapted to this new niche, feeding on human and domestic animals' blood as a zoonosis. Trypanosoma cruzi, their reservoirs, and vectors have existed in nature for millions of years. The primitive trypanosomes were monogenetic parasites of insects that were not bloodsuckers. When these insects acquired the habit of blood sucking, the trypanosomes underwent morphological and functional changes such that they developed undulating membranes and flagella so that they could circulate in the blood of vertebrates (Hoare 1972). Triatomines have been known since the XVI century (Lent & Wygodzinky 1979), but their adaptation to home environments is more recent. Although human infection in 9000-year-old mummies is known (Afderheide et al. 2004), endemic Chagas disease has become established through the forest clearance due to human action over the last two to three centuries (Aragão 1983). It is estimated that 15 to 16 million people are infected with T. cruzi in Latin America, and 75 to 90 million people are exposed to infection. Determinants of Chagas infection Chagas infection, which was initially a wild enzootic disease, was transformed into an anthropozoonosis when man invaded the wild ecotope and occupied the physical space through his predatory action of clearing the forests, displacing the wild animals, and constructing houses and outhouses that the triatomines could easily adapt to. Thus, triatomines began to use man as a food source, and three intercommunicating cycles became established: the wild, domestic surrounds, and domestic cycles. Some wild animals like the opossum, one of the longest known reservoirs of T. cruzi, may have an important role in the epidemiology of Chagas infection, since this mammal circulates from the wild environment to areas around homes and even into homes in search of food. As has been demonstrated, these animals not only carry the trypomastigote form of T. cruzi in their bloodstream but also house all forms of the parasite in their scent glands, and eliminate it from there (Deane et al. 1984). Conversely, domestic animals like cats and dogs may invade the wild environment to hunt, thereby becoming infected orally and bringing the infection into the home and areas surrounding the home. The adaptation of triatomines to human homes, along with the circulation of T. cruzi among them and wild and domestic animals, is certainly the most important determinant for establishing human infection. Although there has been speculation regarding a genetic transformation leading to this adaptation, it seems primarily to be the result from the search by triatomines for a new food source when the original source became scarcer. Forattini (1980) put hencoops into various ecotopes outside of domestic environments and clearly demonstrated this adaptation. The mechanisms for the adaptation of wild triatomines to homes and areas surrounding homes have not been fully clarified. Democritus' saying is fitting here, as so well used in the classic book by Jacques Monod "Le hasard et la nécessité" (Monod 1970): "everything that exists in the Universe results from chance or necessity". In fact, the mechanism for the adaptation of triatomines to human homes seems to fit in well with this concept: the notion of chance, when wild triatomines are passively brought into the home and areas surrounding the home with the wood for construction and fuel and the palm leaves for roofing houses and outhouses, or when they are attracted by the light; or the notion of need, when man invades the wild ecotope, clears the forest, contracts his houses and displaces the animals, thereby obliging triatomines to seek "artificial niches" in homes and areas surrounding homes, in search of alternative food sources among humans and domestic animals. Some species like Triatoma infestans are today totally domesticated, except in Bolivia, while others like Panstrongylus megistus, T. brasiliensis, T. sordida and T. pseudomaculata in Brazil are ubiquitous: sometimes wild and sometimes domesticated or semi-domesticated. Fig. 1 shows palm tree habitat of opossum and triatomines infecteded with T. cruzi invading huts. For human Chagas infection to occur under natural conditions, first of all there needs to be contact between man and infected triatomines. For this, there is a series of variables regulating the chances of infection, among which the following can be highlighted: infection rates, the time interval between biting and defecation, the number and volume of evacuations per unit of time, the number of parasites eliminated, the percentage of infecting forms and their penetration capacity, the intensity of the pruritus caused during the biting, etc. For example, although some species are domesticated, they live at greater separations from man, in roof spaces and cellars, like T. rubrofasciata, and feed on domestic animals, particularly rats, and occasionally on humans. Other with a higher degree of anthropophilia lives in closer proximity, for example in the bedroom. The rates of natural infection of triatomines caused by T. cruzi vary enormously with the species and the closeness of its contact with the reservoirs for the parasite. Among the domesticated triatomines, usually not more than 5% are infected. There are also large differences in the time interval between biting and evacuation according to species, sex, and evolutive stage of the insect. The pioneering investigations on North American (Wood 1951) and South American (Dias 1956, Zeledon et al. 1977) triatomines found that the percentage of Rhodnius prolixus that evacuated just after or while sucking ranged from 40 to 62.5%, T. infestans from 10 to 54.4%, R. neglectus from 12 to 50%, P. megistus from zero among males to 40% among females, T. sordida from zero among nymphs to 23.1% among male adults, and finally none of the nymphs of T. vitticeps. In an interesting experimental study carried out in our laboratory (Borges-Pereira et al. 1988), a mean of 140 parasites per defecation during feeding was found among eight triatomine species infected with T. cruzi. The percentage of positive defecations was 55.1% (mean of 232 parasites) for P. megistus, 52.9% (128 parasites) for R. prolixus, 47.1% (76 parasites) for T. vitticeps, 44.4% (106 parasites) for T. infestans, and 29.4% (98 and 51 parasites respectively) for T. sordida and T. pseudomaculata. This indicated that P. megistus, R. neglectus, R. prolixus and T. infestans had high vector efficiency, while the other species studied had lower efficiency. In an exhaustive and detailed experimental study seeking the best insect for xenodiagnosis, among nine triatomine species, it was found that P. megistus was the most efficient for such tests. Significant differences between the percentages of the forms of T. cruzi in the intestines of the different species were also found (Szumlewicz & Müller 1979, 1982, 1987). On the other hand, it was demonstrated (Soares & Marsden 1986) that T. cruzi was only capable of penetrating through the bite site in 24% of the animals (mice). It could finally be concluded that the chances of natural infection of humans by T. cruzi would not be very high if it were not for the long and intense contact with infected triatomines that is accumulated over the course of life, with hundreds of opportunities, from which one such contact might lead to infection. Other infection mechanisms such as blood transfusion, consumption of infected material (including breast milk), and accidental infection in laboratories will not be discussed here because of space requirements. The mechanisms for transmitting Chagas infection can be summarized as follows (Coura et al. 2007): Principal mechanisms
Secondary mechanisms
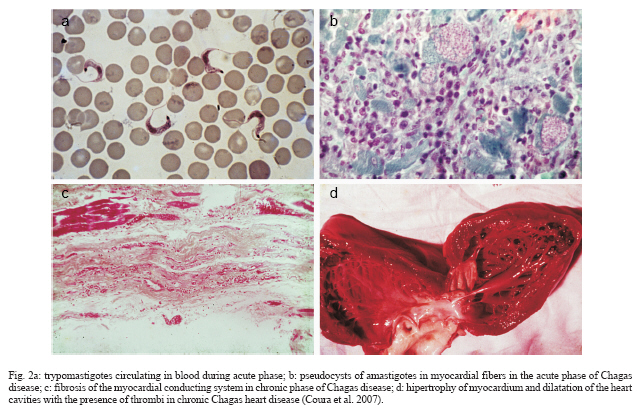
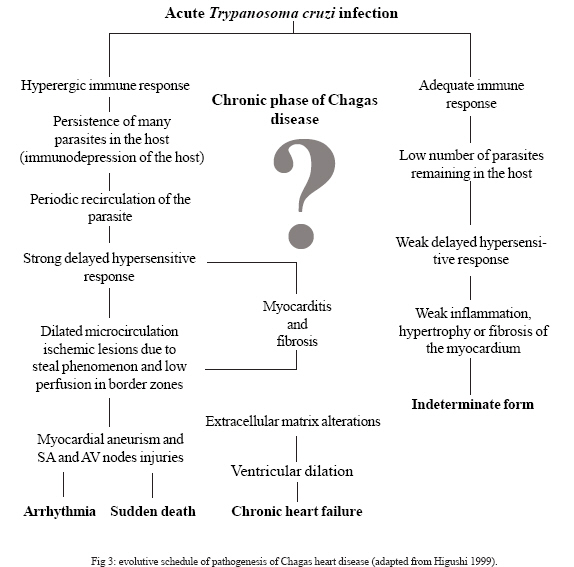
Determinants of Chagas disease The determinants of Chagas disease result from the quantity of parasites in the initial infection; the infecting forms in the initial inoculum (number of trypomastigotes); the lineage of T. cruzi inoculated (I, II, Z3 or hybrid Z1/Z3); reinfection; the quality of the strains and clones (biodema); the specific clonal-histotropic receptors of the host; and the patient's initial and late immune response (Coura 1988, Macedo & Pena 1998, Andrade et al. 2006, Teixeira et al. 2006). The parasites deposited on skin wounds or mucosa stimulate a local inflammatory reaction (inoculation chagoma or Romaña's sign) with a lymphoreticular response. The circulating trypomastigotes enveloped by macrophages are taken to the liver, spleen, lymphatic ganglia, and skeletal and cardiac muscles to form pseudocysts of amastigotes. With the rupture of the pseudocysts in the myocardium or myenteric plexuses, acute myocarditis mediated by TCD4+ and TCD8+ cells and interleukins (mainly IL-2 and IL-4) occurs. The inflammatory reaction leads to muscle and neuron cell destruction, which is maintained by the presence of T. cruzi or its fragments and by the DNA of the parasite, with a late hypersensitivity reaction, dilatation of the microcirculation and fibrosis, thus inducing dilated chronic myocardiopathy, arrhythmia, dysperistalsis, megaesophagus, and megacolon. Fig. 2 shows T. cruzi and the lesions that it causes in the heart. Three characteristics of the presentation of Chagas disease need to be discussed here: the acute or initial phase; the chronic phase with its indeterminate, cardiac, and digestive (enlarged) forms; and the congenital form, which presents particular characteristics. Among the determinants of Chagas disease, the following need to be considered: the inoculum of T. cruzi in the initial infection and the opportunities for reinfection; the biological characteristics of the infecting strains and clones, and particularly their histotropism; and the host response at cell and humoral level. The inoculum in the initial infection is expected to be a factor of great importance in the development of Chagas disease. All the indications from experimental studies are that this inoculum is generally small. In one experimental study (Borges-Pereira et al. 1988), a mean range of 51 to 276 T. cruzi per defecation was found in a study on eight species of infected triatomines, although sometimes single defecations containing more than 1500 parasites were found. Even so, the natural inoculum is infinitely smaller than what is used experimentally on laboratory animals, which reaches 10,000 parasites or more for one mouse. The vast majority of cases of the acute or initial form of Chagas disease in Brazil present few or no symptoms, possibly because of the small inoculum. Among 510 chronic cases in several Brazilian states that we followed up over the last 30 years, we found that less than 1% had a history of an acute phase (Coura et al. 1983). A study following up 544 individuals who were exposed to natural infection by T. cruzi over a 16-month period, it was found that 14 (2.57%) became infected, of whom only one third presented symptoms compatible with the acute phase of the disease (Teixeira 1997). On the other hand, a study covering almost three decades of following up cases with a known acute phase, in the municipality of Bambuí, found that the most severe chronic cases originated from cases that have had a severe acute form. From this, it could be inferred that the initial inoculum and/or the infecting strain of T. cruzi had an influence on the evolution of the disease (Dias 1982). Reinfection by T. cruzi is expected to be rare because of the concomitant immunity induced by the primary infection. Nonetheless, its existence has already been proven both experimentally and in human cases. A wide-ranging study on the "influence of exposure to reinfection on the evolution of Chagas disease" has even described a case of proven reinfection that led to death during the acute phase (Macedo 1976). The biological characteristics of the strains and clones of T. cruzi, and particularly its tissue tropism, certainly have an important function as determinants of Chagas disease and its clinical form. Different strains of T. cruzi have been grouped according to their biological characteristics and cell tropism in mice, to create three characteristic groups (Andrade 1976). In another study, it was found that clones from the same strain produced lesions of differing intensities experimentally (Postan et al. 1983). In a study under our guidance, it was seen that there was a lack of correlation between experimental histopathological findings from mice and 17 strains of T. cruzi that were isolated from patients with different clinical forms of human Chagas disease (Schlemper et al. 1983). Two pathogenic mechanisms are known for infection by T. cruzi: the first, described by Gaspar Vianna in 1911, consists of a local inflammatory reaction with necrosis, tissue destruction, and scar formation with fibrosis; the second, which is more complex to understand and prove, is the immunological mechanism or mechanisms (Teixeira et al. 1975, Andrade 1999). However, this auto-immunity is insufficient to explain all the pathogenesis of Chagas disease (Tarleton 2003). Sensitization of TCD4+ and TCD8+ lymphocytes by T. cruzi, with the development of anti-myocardial cells, associated with migration and activation of macrophages and the release of platelet aggregation factors, thereby respectively inducing chronic Chagas myocarditis and myocardial ischemic lesions, may explain the findings encountered in cases of chronic Chagas cardiopathy (Higushi 1999). On the other hand, the neuron destruction in the heart, esophagus, colon, and other hollow viscera may be explained both by direct inflammatory phenomena and by immunological mechanisms that result in the cardiopathy, megaesophagus, megacolon, and other enlargement phenomena seen in Chagas disease (Prata 2001, Teixeira et al. 2006). The lesions of the acute phase of the disease are characterized by the presence of localized inflammatory reactions, with predominance of mononuclear cells at the foci of the pseudocyst ruptures, occasionally with the formation of granulomas located mainly in muscle and cardiac tissue. In certain cases, there may be lymphoreticular hyperplasia in the lymphatic ganglia, liver, and spleen and the presence of macrophages invaded by parasitic cells. In other cases, or concomitantly, there may be diffuse acute myocarditis, with interstitial edema, hypertrophy of myocardial fibers, and dilatation of cardiac cavities. The destruction of the cardiac neurons and myenteric plexuses, with reductions in the numbers of neurons, begins in the acute phase and continues in the chronic phase of the disease (Koberle 1961, Andrade 2000, Andrade 2005) (Fig. 3: evolutive schedule of Chagas heart disease; adapted from Higushi 1999). The indeterminate chronic phase (asymptomatic) has practically no anatomopathological translation, except for occasional isolated inflammatory foci in the myocardium and a limited reduction in the numbers of cardiac neurons and myenteric plexuses that is insufficient to produce clinical manifestations (Andrade et al. 1997). On the other hand, in the cardiac form of the chronic phase, there may be extensive myocardial fibrosis, destruction of the conduction system, and a large reduction in the numbers of cardiac neurons. It is worth pointing out that, in these cases, isolated foci of acute inflammatory reaction are occasionally present, as if this were a reactivation of the process. In chronic Chagas cardiopathy cases, it is common to find hypertrophy of myocardial fibers and dilatation of the cavities, with the presence of thrombi, fibrosis, and thinning of the ventricular apices, particularly in the left ventricle. These are sometimes characterized as true apical aneurysms and are almost always internally carpeted with organized blood thrombi. Also in the chronic phase, there is frequently a large reduction in the number of neurons in the myenteric plexuses, particularly in the esophagus and colon, which leads to dysperistalsis and dilatation of these organs that is characterized by megaesophagus, megacolon, and other enlargements of hollow viscera such as the bladder, ureter, gallbladder, and other less common examples (Andrade & Andrade 1966, Lopes & Chapadeiro 1997, Dias & Macedo 2005). The congenital form of Chagas disease seems to occur solely in pregnant women who have a lesion in the placenta that favors penetration by T. cruzi as far as the chorionic villi, where amastigote forms multiply (probably in Hofbauer cells) and subsequently invade the fetal circulation (Bittencourt 1963, Carlier & Torrico 2005). Clinical phases and forms: morbidity-mortality Chagas disease presents an initial or acute phase with evident parasitemia seen in direct examination of the blood. In most cases there are no symptoms, but in symptomatic cases there are entry point signs (inoculation chagoma or Romaña's sign (Fig. 4), with fever, generalized adenopathy, edema, hepatosplenomegaly, myocarditis, and meningoencephalitis in severe cases. This is followed by a chronic phase that in most cases presents as an indeterminate form (asymptomatic, with normal results from electrocardiogram and x-ray of the heart, esophagus, and colon), which may evolve to the cardiac or digestive form (megaesophagus and megacolon), or cardiac and digestive forms together. The so-called congenital form mentioned earlier may also occur, by means of transmission across the placenta or through the birth canal during delivery, and this may give rise to abortion, prematurity or organic lesions in the fetus (Bittencourt 1963, Carlier & Torrico 2005). In cases with immunosuppression, the chronic infection may become acute again, thereby producing diffuse myocarditis, lesions of the central nervous system, and severe meningoencephalitis. The clinical phases and forms of Chagas disease can be summarized as follows (Coura et al. 2007): Acute phase
Chronic phase
Congenital form
Forms in immunosuppressed individuals
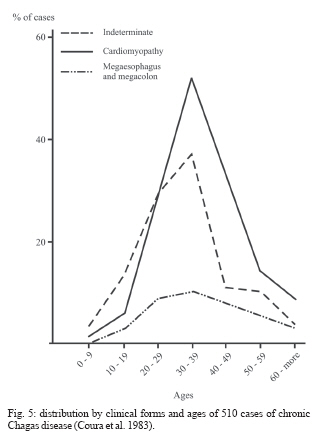
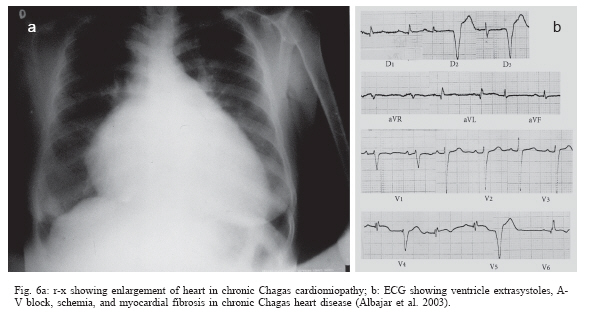
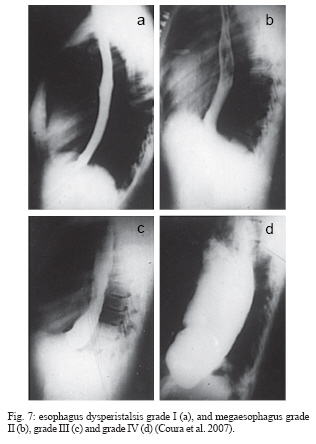
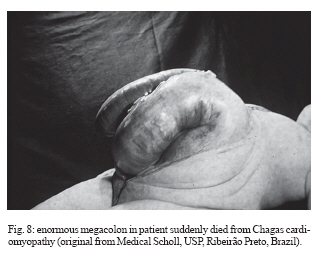
In the chronic phase, Chagas infection may present as an indeterminate form, in which approximately 40% of the infected individuals remain totally asymptomatic, with anatomically and physiologically normal x-ray results for the heart, esophagus, and colon and no changes seen on electrocardiograms. Although they are asymptomatic, these patients present positive serological reactions for T. cruzi infection and, for many of these patients, the xenodiagnosis and PCR results may be repeatedly positive for many years. Thus, they display a veritable equilibrium between the parasite and host. There is great regional variation in the morbidity of Chagas disease. The indeterminate form can vary from 40 to 90% of the cases (Coura et al. 1985, 1999) (Fig. 5). The chronic cardiac form is the most expressive manifestation of Chagas disease, both because of its frequency and because of its severity. It generally appears between the second and fourth decades of life, five to fifteen years after the initial infection. The signs and symptoms of chronic Chagas cardiopathy result from arrhythmia, cardiac insufficiency, auricular-ventricular and branch blockages, and thromboembolism. In our first series of 100 cases studied (Coura 1965), we found the following manifestations: effort dyspnea 65%, palpitations 54%, extrasystole 51%, apical systolic murmur 47%, vertigo and/or fainting 37%, lower-limb edema 35%, precordial pain 37%, double second heart sound at the pulmonary focus 24%, hepatomegaly 19%, double first heart sound at the mitral focus 18%, bradycardia 17%, tachycardia 15%, hypophonesis of the heart sounds 10%, gallop rhythm 6%, anasarca 6%, and convulsions 5%. Fig. 6 shows the increased area of the heart and the electrocardiographic abnormalities in chronic Chagas cardiopathy. In the chronic digestive form of the disease, the clinical manifestations result from dysperistalsis of the esophagus and colon caused by destruction of the myenteric plexuses, which consequently leads to megaesophagus and megacolon. Although isolated cases of autonomic disorders of the esophagus have been described in the acute phase of the disease, these occur mostly in the chronic phase, when the dysperistalsis and spasms are accompanied by enlargement of the esophagus. Fig. 7 shows the evolution of megaesophagus and Fig. 8 the megacolon. The prognosis for Chagas disease depends on the clinical form and the complications during its evolution. In the acute phase, it depends on the patient's age and the severity and location of the lesions. In general, the acute phase is very severe among children less than two years old, and it is almost always fatal in those with myocarditis, cardiac insufficiency, and meningoencephalitis. The prognosis may also be very poor in the congenital form, which may lead not only to abortion and prematurity but also to organic lesions in the liver, spleen, heart, and central nervous system, with neurological sequelae and mental deficiency. Many cases may be asymptomatic and remain in the indeterminate form. In the chronic cardiac form, the prognosis varies considerably from one case to another. Patients with minimal lesions such as blockage of the right branch alone or unifocal ventricular or auricular extrasystole tend to remain stable and most of them survive for long periods and often end up dying for other reasons. Patients with complex arrhythmia, multifocal extrasystole, paroxystic tachycardia, auricular fibrillation, third-degree A-V blockage or cardiac insufficiency have a very poor prognosis. A third group of patients with slightly increased heart area and changeable electrocardiographic findings and clinical manifestations have an uncertain prognosis (Nogueira & Coura 1990). Studies carried out by our group (Abreu 1977, Coura et al. 1985, Borges Pereira et al. 1985) in field areas in Minas Gerais have shown that the mortality due to Chagas cardiopathy increases progressively from 30 to 59 years of age. In these studies, we showed that the death rate due to Chagas cardiopathy was 8.9% among patients followed up for six years and 17% in another group followed up for ten years. These same authors found that sudden death occurred in two thirds of the cases, while the other third died because of cardiac insufficiency. The prognosis for the digestive and indeterminate forms is generally good, except in cases of the digestive form with complications (esophageal cancer, obstruction with twisting, and colon necrosis). None of the cases of the digestive or indeterminate form that we observed over six and ten-year periods progressed to death (Coura & Borges Pereira 1984). Chagas disease in the Amazon region Several acute cases of human Chagas disease have been reported from the Amazon region most of them by T. curzi I, Z3, and hibrid ZI/Z3. In the localities where this disease has been reported, the chronic form of this disease is considered to present low endemicity. The first acute cases in the Amazon region were reported by Floch and Tasque (1941) and by Floch and Camain (1948) from French Guiana. Shaw et al. (1969) described another four acute cases in Belém, the capital of the state of Pará, in Northern Brazil. Since then, more than four hundred acute cases have been reported, most of them from outbreaks probably by oral transmission in the states of Pará, Amapá, and Amazonas, Brazil (Valente &Valente 1993, Valente et al. 1999, Pinto et al 2004, Coura 2006). Serological surveys and cross-sectional studies carried out by Fundação Nacional de Saúde from 1975 to 1980 in different states in the Brazilian Amazon region and by Coura et al. from 1971 to 2002 in the state of Amazonas showed prevalences ranging from 2.4% to 13.2% (Camargo et al. 1984, Coura et al. 1999, 2002). Sporadic chronic cases of Chagas disease have also been reported from the Brazilian Amazon region (Albajar et al. 2003, Junqueira et al. 2005, Xavier et al. 2006). The ten top Chagas disease needs 1. To clarify the pathogenesis and evolutive pattern of Chagas disease. 2. To develop an effective drug for the acute and chronic phases. 3. To improve a specific and fast test for T. cruzi I, II, Z3, and Z1/Z3 infections. 4. To standardize the techniques for serological diagnosis and control of cure evaluation. 5. To compare the morbidity induced by the four types of T. cruzi. 6. To study the metacyclogenesis of different strains of T. cruzi. 7. To analyze the risk that wild triatomines may become adapted to human dwellings. 8. To evaluate the morbidity of Chagas disease in the Amazon region. 9. To establish a surveillance and control program for Chagas disease in the Amazon region. 10. To prioritize Chagas disease control programs in Latin America and Mexico. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07142f8.jpg] [oc07142f7.jpg] [oc07142f2.jpg] [oc07142f6.jpg] [oc07142f1.jpg] [oc07142f5.jpg] [oc07142f4.jpg] [oc07142f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}