 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
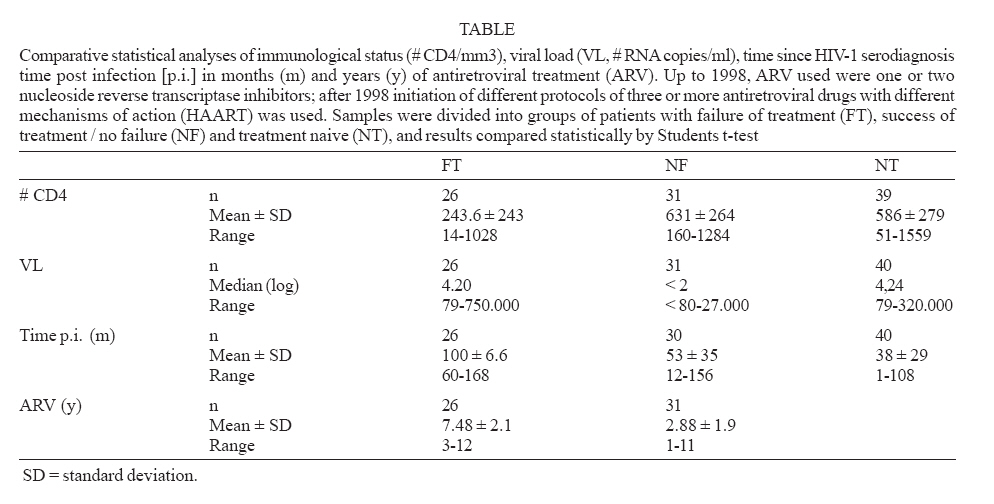
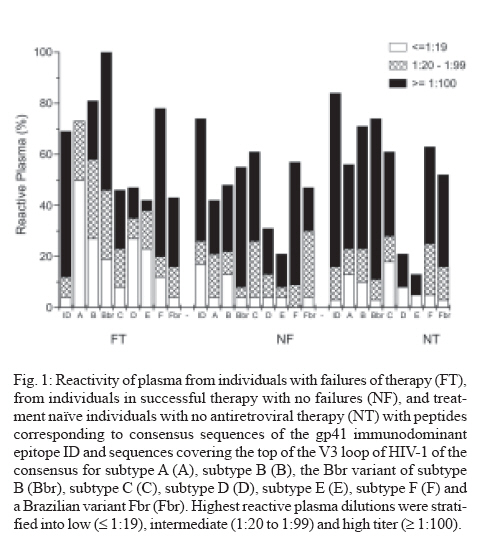
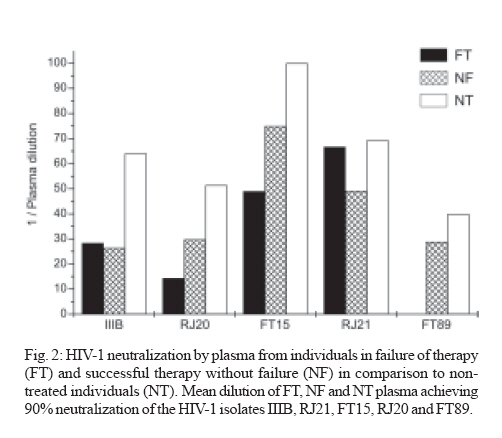
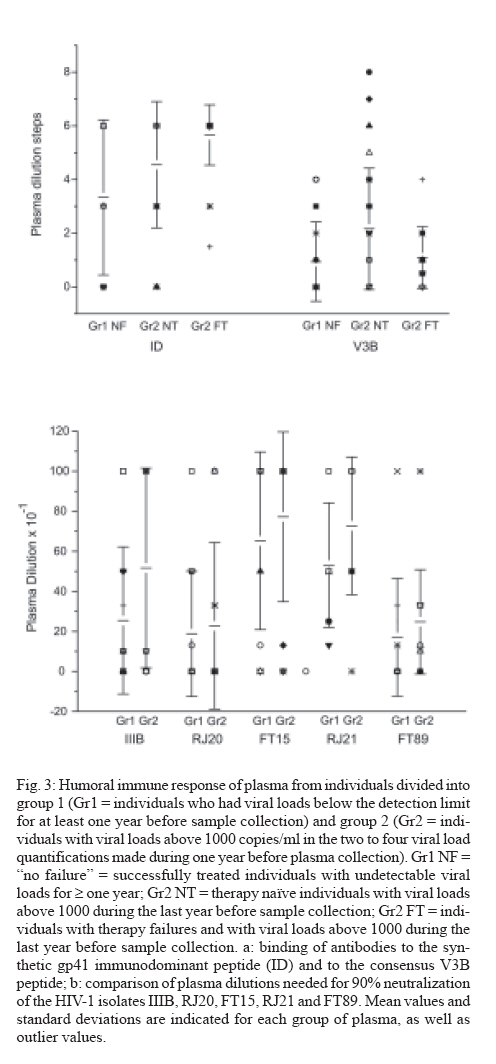
Memórias do Instituto Oswaldo Cruz, Vol. 102, No. 7, November, 2007, pp. 817-825 Anti-human immunodeficiency virus type 1 humoral immune response and highly active antiretroviral treatment Vera Bongertz/+, E Priscilla Ouverney, Saada Chequer Fernandez, Beatriz Grinsztejn*, Valdilea Veloso*, José C Couto-Fernandez, José H Pilotto*, Mariza G Morgado Laboratório
de AIDS e Imunologia Molecular, Instituto Oswaldo Cruz-Fiocruz,
Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil *Instituto
de Pesquisa Clínica Evandro Chagas-Fiocruz, Rio de Janeiro,
RJ, Brasil Financial support: PAPES III - Fiocruz Received
28 March 2007 Code Number: oc07148 Highly active antiretroviral treatment (HAART) of human immunodeficiency type 1 (HIV-1) infection is very effective in controlling infection, but elimination of viral infection has not been achieved as yet, and upon treatment interruption an immediate rebound of viremia is observed. A combination of HAART with an immune stimulation might allow treatment interruption without this rebounding viremia, as the very low viremias observed with successful HAART may be insufficient to permit maintenance of a specific anti-HIV-1 immune response. The objective of this study was to compare the humoral immune response of individuals undergoing successful HAART (NF=no failure) with that of individuals with evidence of failure of therapy (FT) and to verify if the viremia peaks observed in individuals with therapy failure would act as a specific stimulus for the humoral anti-HIV-1 immune response. Antibodies binding to gp120 V3 genotype consensus peptides were more frequently observed for FT, mainly against peptides corresponding to sequences of genotypes prevalent in the Rio de Janeiro city area, B and F. HIV-1 neutralization of HIV-1 IIIB and of four primary isolates from Rio de Janeiro was less frequently observed for plasma from the NF than the FT group, but this difference was more expressive when plasma from individuals with detectable viremia were compared to that of individuals with undetectable viral loads in the year before sample collection. Although statistically significant differences were observed only in some specific comparisons, the study indicates that presence of detectable viremia may contribute to the maintenance of a specific anti-HIV-1 humoral immune response. Key words: antibodies - seroreactivity - neutralization - antiretroviral treatment - treatment failure Anti-HIV highly active antiretroviral treatment (HAART) reduces HIV in peripheral blood and reverts the characteristic immunodeficiency, and has benefited patients increasing individual survival at least 13-14 years (Vermund et al. 2006). However, no elimination of viral infection has been achieved, and the antiretroviral drugs will probably have to be taken throughout the life of the patient. Attempts to stimulate a potent specific anti-HIV-1 immune response to permit control of viral replication. After HAART has reduced viremia to undetectable levels, have so far been less than successful. During successful HAART, a decline in antibody response is generally observed (Fournier et al. 2002, Devito et al. 2006), with cases of seroreversion being described (Amor et al. 2006) and the extent of immune reconstitution in HAART treated individuals is controversial, apparently depending on the immune status of the individuals at the start of therapy (Cheonis et al. 2005). However, the benefits of HAART are evident even when treatment apparently fails, leading neither to a complete control of viremia nor to a complete reconstitution in CD4 T lymphocytes (Brígido et al. 2004, Kovacs et al. 2005). During successful anti-HIV therapy, a reduction in virus specific cellular immunity occurs (Dalod et al. 1998), possibly due to inefficient CD8 T lymphocyte stimulation at the consequent very low antigenic concentrations (Bucy et al. 2001). It is also thought that the low level of HIV replication during successful HAART may prevent induction of a high titer antibody response (Popovic et al. 2005, Bailey et al. 2006), and, although sporadic cases of a high autologous neutralizing antibody (NAb) have been reported, the NAb response during HAART is poorly understood (Kimura et al. 2002, Dreyer et al. 1999). Titers of "binding antibodies" (bAb = antibodies detectable through their in vitro binding to synthetic proteins or peptides) have been shown to decrease during successful HAART (Morris et al. 2001, Bailey et al. 2006). However, generation of specific anti-HIV antibodies in absence of detectable viremia has been described, indicative of retention of HIV antigen over long periods (Kim et al. 2001, Popovic et al. 2005) and restoration of normal B cell functions during HAART (Morris et al. 1998, David et al. 1999). Nevertheless, there are indications that the moment of HAART initiation defines the consequential restoration in the immune response (Mocroft et al. 2003, Manzardo et al. 2007, Chehimi et al. 2007): the extent of immune response alterations caused in HIV infection will define restoration, but the quantitative immune stimulation induced by HIV will also be important for the extent of specific anti-HIV response detectable during or after successful antire-troviral therapy. Intermittent structured treatment interruptions may stimulate antiviral immunity through an autoimmunization process (Jacobson et al. 2006), although this process seems to be controversial (Ioannidis et al. 2000). However, a stimulation of the CD8 T lymphocyte anti-nef response has been shown to occur (Benito et al. 2006). Also, some studies indicate that occasions of viral rebound or "blips" may benefit patients by increasing their specific immune response (Kimura et al. 2002, Sullivan et al. 2003, Karlsson et al. 2004, Macias et al. 2005), including the titers of bAb (Binley et al. 2000, Killian et al. 2006) and the NAb response (Binley et al. 2000, Montefiori et al. 2001, Kimura et al. 2002, Wang et al. 2002). It is known that, depending on the drug treatment of the transmitting individuals, up to 20% of new patients were infected with strains that were already drug resistant (UK Collaborative Group on HIV Drug Resistance 2007). Resistance data are available for some countries, with rates increasing worldwide since 1997. Despite the large use of HAART-based regimens in the treatment of HIV/AIDS patients in Brazil, the rate of primary HIV drug resistance is very low when compared with that of developed countries (Soares et al. 2004). The present study was carried out in an attempt to evaluate if an effective autoimmunization occurs with a stimulation of anti-HIV-1 humoral immune response in individuals who display peaks of viral load resulting from therapy failure, whether induced by lack of adherence to therapy schedules or by the appearance of drug resistant viral quasispecies. The drug-resistance mutations were evaluated in all patients of this study under treatment with HAART, through the genotyping of the HIV pol gene (Eyer-Silva et al. 2006). SUBJECTS, MATERIALS AND METHODS Plasma Samples - HIV-1 infected individuals followed at the Evandro Chagas Clinical Research Institute of Fiocruz between april 2003 and october 2004 were invited to participate in this study, and a total of 97 agreed in participating, after signing the informed consent form (project approved by the Fiocruz Ethical Committee n. 172/02). Results obtained from individuals selected to participate, but who did not meet the criteria defined for FT (failure of therapy), NF (no failure) or NT (no therapy) were excluded from the data shown. All patients had at least 4 medical visits, with lymphocyte typing (FACScan, Becton-Dickinson, San José, CA) and viral load determinations (nucleic acid sequence-based amplification NASBA, NucliSens, Organon Teknika, Boxtel, Netherlands). A total of 26 patients had been for at least one year in successful therapy, as indicated by decreasing viral loads and stable or increasing concentrations of CD4 lymphocytes in peripheral blood (group NF), and had no record of any therapy failure events or therapy interruptions. A group of 31 individuals in therapy with episodes of failure of therapy (FT), defined as patients experiencing virologic failure (sudden rises of HIV RNA to more than 5,000 copies RNA/ml, caused either by lack of adherence and/or viral resistance to antiretro-virals), called group FT, was included. As an outside control group, a total of 40 antiretroviral naive individuals (NT) were included. Table indicates mean values for information collected for the different groups. All patients were followed according to the Brazilian Ministry of Health HIV/ AIDS guidelines, receiving free antiretroviral therapy when numbers of circulating CD4 lymphocytes dropped below 350/ mm3, viral loads were greater than 30,000 copies HIV-1 RNA/ml in peripheral blood or with viral loads increasing by more than one log in three months. Accordingly, all individuals receiving HAART agreed to collect four viral load and CD4 lymphocyte determinations per year and for evaluation of drug resistance mutation profiles. Seroreactivity - Several custom made synthetic peptides (Chiron Mimotopes, San Diego, CA, USA) were used: a) corresponding to the consensus sequences of the gp120 V3 loop of HIV-1 subtypes: A (KSVHIGP-GQAFYAT), B (NTRKSIHIGPGRAFY), Brazilian Bbr (NTRKSIHMGWGRAFY), C (KSIRIGPGQTFYAT), D (RQRTHIGPGQALYTT), E (RTSITIGPGQVFFYR), F (RKSIHLGPQAFYTT), and of a representative Brazilian F isolate (FBR = NTRKSIPLGPGRAFY), and b) to the immunodominant epitope in gp41 of the HIV-1 isolate MN (ID = GFWGCSGKLICTTTVPWNAS). Duplicates of sequentially diluted (1:100 at a first step, followed by serial dilutions using factor 2) heat inactivated plasma were incubated (1h/37oC followed by an overnight incubation at 4oC) with different biotinylated synthetic peptides (5 mg/well) bound to multiwell plates (Maxisorb, Nunc, Roskilde, Denmark) precoated with streptavidin (10 mg/well). Specific reactivity was assessed (including two 8M urea washes after antibody incubation according to Casseb et al. 2002) by peroxidase-conjugated anti-human-IgG binding and peroxide/ tetrametilbenzidine revelation. Reactivity was assessed as dilution titer reaching at least twice the mean binding of the duplicate control plasma pools (obtained from local HIV negative blood donors) included in each assay plate. Absorbency was measured at 450 nm. For some of the samples tested in duplicate, reactivity with the peptides was discordant even after repetition, probably indicating low affinity binding partially removed by the 8M urea washes. These results were considered doubtful and not included in this report, leading to differences in numbers of samples tested with the different synthetic peptides (Bongertz et al. 2003). Neutralizing Antibodies - Individual heat inactivated plasma from the different groups were tested as to their capacity to neutralize the reference HIV-1 isolate IIIB (kindly donated by Dr EM Fenyoe, Lund University, Sweden), adapted to replication in lymphocytic cells lines expressing CD4. For use in this study, viral stocks from two replication cycles in pre-activated normal human peripheral blood mononuclear cells (PHA-PBMC NHu, mixture of at least two donors) were pooled. First to third passage viral stocks of the primary HIV-1 isolates FT015 (subtype F, in this report called FT15), FT089 (subtype B, in this report called FT89), and third to fifth passage of 95BRRJ021 (subtype F, in this report called RJ21) and 95BRRJ020 (subtype B, in this report called RJ20) were also included (Bongertz et al. 2000). At least three 3-fold plasma dilutions, using 1:10 as the first step, were used. Neutralization of the HIV-1 isolates was tested using PHA-PBMC NHu (105 cells/well), using a multiplicity of infection (MOI) of 0.001-0.005 (10-50 infective units per well), according to the WHO-UNAIDS Guidelines (WHO-UNAIDS 2002). Quantification of the HIV-1 p24 antigen (HIV-1 p24 ELISA Zeptometrix Corporation, Buffalo, NY) was carried out on the seventh day of cell culture. Positive control wells containing virus and cells (without human serum), assay wells containing virus, cells and serially diluted plasma, negative control wells (containing virus, cells and normal human serum), background values (wells containing only cells) and wells used for determining the tissue culture infective dose 50% (TCID50%) of the virus isolate were prepared simultaneously (Bongertz et al. 2001). Approximate plasma concentrations needed for neutralizing 90% and 50% of the viral input (90% or 50% neutralization levels, respectively) were derived from linear regression curves (MicroCal Origin Software Inc, Northampton, MA, USA) or directly from the neutralization curve. HIV-1 subtyping and drug resistance genotyping - Genomic DNA was extracted from 200 ml of whole blood from all 97 individuals included in this study using the QIAamp DNA kit (Qiagen Inc, Chatsworth, CA, USA), according to the manufacturer's protocol. The HIV-1 genetic subtype determination was performed analyzing the envelope (env) C2-V3 region of gp120, amplified by nested PCR as previously described (Guimaraes et al. 2002). The presence of drug resistance mutations to nucleoside reverse transcriptase inhibitors (NRTI), non-nucleoside RTI (NNRTI) and protease inhibitors (PI) was performed targeting two genomic regions of the polimerase (pol) gene, amplified by nested PCR, covering the whole protease (codons 1-99) and the first 235 codons of RT as described elsewhere (Eyer-Silva et al. 2006). All PCR products were purified using the Qiagen PCR purification kit (Qiagen) according to the manufacturer's protocol, sequenced using the ABI PRISM Big Dye Terminator Cycle Sequencing Ready Reaction Kit v.3.1 and analyzed in a ABI 3100 automate sequencer (Applied Biosystem, Foster City, CA, USA). Resistance mutations and phylogenetic analyses - Sequence chromatograms were aligned in PC/Windows using SeqMan software (DNAStar, Madison, WI) and manually edited. Edited sequences were then aligned to reference sequences representative of all HIV-1 subtypes obtained from the Los Alamos database (http://hiv-web.lanl.gov) in ClustalW (Thompson et al. 1994). Aligned sequences from env and pol genes were subjected to phylogenetic inference through the neighbor-joining method and Kimura 2-parameter model of the MEGA 2.1 package (Kumar et al. 2001) for assignment of HIV-1 subtype. The genotypic interpretation of antiretroviral drug resistant mutations in PR and RT was carried out by electronic submission to the Stanford database (http://hivdb.stanford.edu). Distinction of primary and secondary HIV-1 drug resistance mutations was based on the International AIDS Society guidelines (htpp://iausa.org.us). Statistical analyses - For evaluation of the results obtained the Instat software (Graph Instat tm, 1990-1994, San Diego, CA, USA) was employed, using the ANOVA non-parametric test, unpaired t-student test for comparison between ages, CD4 T cell counts, and viral loads; and contingency Table (chi-square or exact Fisher test depending on sample numbers) for categorical variables. Linear regression was plotted using the Micro Cal Origin software (micro Cal Origin, Northampton, MA, USA, 1991-1992). RESULTS Plasma Samples - All groups had a slightly higher participation of males and the majority indicated that infection occurred by sexual exposure, while ages varied from 18 to 60. However, no statistically significant differences between groups were detected (data not shown). A summary of comparative CD4 lymphocyte numbers and viral loads at sample collection is shown in Table, indicating significantly lower counts of CD4 T lymphocytes (in comparison to the successfully treated and to the non-treated groups, p < 0.0001) and higher viral loads in the FT group (in comparison to the successfully treated group p = 0.0002 and with the untreated control group p = 0.0008). Individuals had been infected for one to 12 years. In the successfully treated NF (p < 0.0001) and the NT control group (p < 0.0001) a significantly higher number of individuals with periods since positive anti-HIV-1 serology of less than two years was identified in comparison to the FT group. Duration of antiretroviral treatment was much longer for group FT than for group NF (p < 0.0001). Very few individuals from the FT group had been in treatment for less than three years, and therefore, no meaningful comparisons could be made between FT and NF for individuals treated during similar periods of time. HIV-1 Genetic Subtype - A total of 83 (85,5%) samples could be successfully subtyped analyzing env C2V3 sequences and 82 (84.5%) using PR and RT sequences. A total of 57 samples (69.5 %) showed concordance in subtyping determination using these genomic regions. Overall, the majority of the samples were classified as HIV-1 subtype B (80%), followed by subtype F (14%), BF recombinant forms (5%). HIV-1 subtype C was identified in one patient (1%). No preferential distribution of genetic HIV-1 subtypes could be observed in the different study groups. Drug Resistance Mutations - In the treatment-naïve group (NT) various combinations of secondary (minor) protease mutations at codon positions L63P (23%), M36I (18%), A71V and V77I (9% each) were observed. At baseline, the major mutation Y181C associated to NNRTI and the V118I and F77L secondary mutations associated to NRT inhibitors were observed in one patient each (2,5% each). The development of resistance-related mutations in genotyped samples from treated patients failing HAART (FT) showed that 70% had resistance to NRTI, 55% to the NNRTI and 45% to the protease inhibitors. Overall, a reduction of ARV susceptibility observed was around 40% to one class of drug, 20% to two classes of drugs and 30% against 3 classes of drugs. Only 10% of the genotyped FT samples showed complete susceptibility to all classes of ARV, and only 4 % of the successful therapy group showed reduction of two classes: NRTI and NNRTI drugs. Presence of resistance-related mutations in genotyped samples from NF successfully treated patients showed that 10% of them had reduction of susceptibility to NRTI drugs and 15% to NNRTI antiretrovirals. No associated resistance mutations were observed simultaneously for both classes of ARV drugs. V3-peptide Binding Antibodies - Frequency of reactivity and recognition titers were unexpectedly high for plasma from the FT group. As Fig. 1 shows, reactivity was lower for plasma from individuals undergoing successful therapy (group NF), and higher for FT individuals. However, statistical analyses showed significance for only some peptide recognition frequencies and reactivity titers. Thus, recognition of the reference gp41 immunodominant peptide (ID) was higher for FT than NF plasma (p = 0.0133), with higher titers (p = 0.0411). Frequencies were also higher for FT than NF for the Brazilian consensus sequence V3Bbr (p = 0.0001), and even higher than for the treatment-naïve control group NT (p = 0.0024). Differences in reactivity frequencies were highest for peptides corresponding to consensus sequences of HIV-1 subtypes prevalent in Rio de Janeiro (B, Bbr, F, Fbr), city of residence for all individuals participating in this study. Analyses of the frequency of peptide recognition by plasma from individuals infected with B, Bbr or F HIV-1 showed that B and F plasma recognized V3B and V3F to a similar extent, but Bbr plasma showed a much more specific recognition of V3Bbr than V3B (p = 0.0209), V3F (p = 0.0038) or V3Fbr (p = 0.0005). This finding was also expressed by the higher titers of NT-Bbr plasma reactivity with V3Bbr in comparison to reactivity with V3B (p = 0.007), V3F (p = 0.0010) or V3Fbr (p < 0.0001), with similar results being observed for FT-Bbr plasma, while the number of plasma in the NF-Bbr group was too low to permit comparison within the group. A higher titer of F plasma with the V3F peptide than V3Bbr or V3Fbr was again statistically significant only within the FT group. However, the number of plasma in each group was too small to permit definite conclusions to be drawn. HIV-1 Neutralizing antibodies - A total of 5 HIV-1 isolates were used for the analysis, although not all plasma could be tested against all isolates due to limited amounts of primary HIV-1 isolate stocks available. Unfortunately, only isolates from individuals in the NT group could be obtained with the high titers needed for neutralizing antibody assays. Two primary isolates of the F subtype (FT15 and RJ21) and two isolates of the B subtype (FT89 and RJ20) were employed, as was the reference isolate HIV-1 IIIB (also subtype B, adapted to growth in lymphocytic cell lines although an expansion in human lymphocytes was employed in the assays). Plasma from treated patents, successfully (NF group) or unsuccessfully (FT group), were less frequently efficient in neutralizing the different HIV-1 isolates, mostly in comparison to plasma from treatment naïve individuals (group NT), with statistically significant differences for neutralization of IIIB (90% neutralization levels, p = 0.0167). Mostly, however, statistically significant differences were observed at 50% levels of neutralization of viral input, and at lower plasma dilutions, where the effect of plasmatic antiretroviral drugs can have affected neutralization results. Broadness of the response, defined as neutralizing more than half the isolates analyzed, indicated a more limited cross neutralization for individuals of the group of plasma from patients with undetectable viral load for at least one year. However, only a few plasma from the FT group showed capacity to neutralize 90% of the input of at least two of three primary HIV isolates tested (22%), while the majority of the plasma from the NT group (73%) did neutralize the majority of the HIV-1 isolates tested (NT vs FT, p = 0.0166; NT vs NF, p = 0.0127), even if only at lower plasma dilutions. Fig. 2 indicates the similarity in mean neutralization titers observed for FT and NF plasma, with no important differences or trends noticeable. No data is shown for neutralization of HIV-1 FT89 by FT plasma as too few plasma (only five) from individuals in actual therapy failure were used in the assay, as revealed by the decoding of the plasma samples carried out after the assays were finished, a precaution taken to diminish objective evaluation of the data. Humoral immune response and viral load analyses - Although identification of failure of therapy was based on high viremia peaks during treatment, for some of the patients viral loads (VL) were already quite low during sampling. Therefore, an analysis based on actual viral load during the last year before sampling allowed a division of the patients into two groups, one (group 1, Gr1) who had undetectable viral loads for at least one year before sampling (two to four VL determinations) and one (group 2, Gr2) with viral loads above 1,000 copies/ ml plasma in all evaluations during the last year before sampling. Fig. 3a shows an example of the higher recognition of synthetic peptides observed for plasma from individuals with detectable viral loads (group 2), although statistically significant only for the g41 ID peptide (Gr1 vs Gr2 FT, p = 0.00252; Gr 1 vs Gr2 NT, p = 0.0252). Similarly, neutralization titers in plasma from group 2 individuals were higher than those from group 1, in neutralization of all 5 HIV-1 isolates tested (Fig. 3b). However, these differences had no statistical significance (p > 0.05). Anti-HIV antibodies vs CD4 lymphocyte counts and vs therapy failures - No relationship between number of CD4 T lymphocytes at sample collection and HIV-1 neutralization could be observed. Equally, no correlation between the nadir of CD4 cell concentration and HIV neutralization or reactivity of the plasma with synthetic V3 peptides could be observed, although a slight trend for higher HIV-1 neutralization titers (but not V3 peptide binding antibodies) was observed in individuals with a higher number of therapy failures (p = 0.09) (data not shown). As several studies have shown a stimulation of the immune response by structured therapy interruptions or episodes of viral rebound or "blips" during successful HAART (Binley et al. 2000, Ioannides et al. 2000, Montefiori et al. 2001, Kimura et al. 2002, Wang et al. 2002, Sullivan et al. 2003, Karlson et al. 2004, Macias et al. 2005, Benito et al. 2006, Jacobson et al. 2006, Killian et al. 2006), it could be expected that a similar stimulation might occur during increases in viremia during therapy failure episodes, at least in individuals whose immunodeficiency is not advanced. This could be an explanation for the unexpected good health observed despite treatment failure in many patients (Brígido et al. 2004, Kovacs et al. 2005). The results presented indicate that individuals who have had episodes of therapy failure indeed show higher frequencies and titers of antibodies recognizing synthetic peptides corresponding to the immunodominant epitope in gp41 and peptides corresponding to consensus sequences of the V3 loop of some HIV-1 genetic subtypes. As expected, the main difference was observed for V3 peptides corresponding to HIV-1 genetic subtypes prevalent in the state of Rio de Janeiro, where all patients included resided. As in Rio de Janeiro the prevalent HIV-1 subtypes, B and F, both present the GPGR sequence at the top of the V3 loop, and the top of the less often recognized V3 peptides (A, D and E) present the GPGQ sequence, this amino acid substitution may be of importance for the antibody response, as should be expected by the difference in charge and size of the residual groups of the amino acids arginine and glutamine. However, this does not explain the very high reactivity observed in FT plasma with the F peptide. Among 70 Brazilian subtype F sequenced env (most from the southeastern region of the country), approximately 34% display the GPGQ tetrapeptide at the top of the V3 loop, and the great majority of these, as well as the B, Bbr and Fbr V3 consensus sequences, have a great similarity in amino acid sequence of the V3 loop anterior arm (G Bello, personal communication). The similarity of humoral immune response between B and F subtype infected individuals could again be verified in the study presented, as differences between B and F plasma were very minor. Plasma from HIV-Bbr individuals, on the other hand, showed a significant preferential recognition of the homologous V3Bbr consensus. Unfortunately, no isolate identified as belonging to the Bbr variant of the B subtype could be produced in the titers and quantities necessary for neutralization assays, so the question of the susceptibility of Bbr primary isolates cannot be discussed in this study. However, plasma from HIV-Bbr infected individuals neutralized the B and F HIV-1 isolates tested to a similar extent as the B and F plasma identified. No association between HIV-1 subtype and disease progression could be observed, however, the data reveal an elevated frequency of drug resistance mutations in the protease gene among drug-naïve individuals from Rio de Janeiro city. The profile of drug resistance mutations in the treated groups showed correlation with the therapy employed and no differences were observed between failure and responder groups. The presence of resistance associated mutations against different classes of ARV drugs supports the importance of resistance testing for guidance of therapy. The neutralization analyses carried out indicate that the stimulation of NAb was not as obvious as that of the bAb: HIV-1 neutralization was quite low for the majority of the plasma analyzed, with few plasma neutralizing any of the isolates at dilutions above 1:50, considered to be a dilution where the drugs present in the plasma of the FT and NF individuals would not affect HIV-1 replication in the NAb assay. However, it is very difficult to be sure of this effect in neutralization assays, as of the 51 plasma tested only three were able to neutralize all isolates tested. The other 48 plasma were able to neutralize one or more isolates but showed no neutralization of at least one isolate tested, even at the 1:10 dilution, indicating a lack of interference of the antiretroviral drugs taken. Agreeing with results obtained before (Bongertz et al. 2005), the F subtype primary HIV-1 isolates were significantly more susceptible to neutralization than the B subtype primary isolates, and even more susceptible than the T cell line adapted isolate HIV-1 IIIB. Broadness of the neutralizing antibody response was highest for plasma from treatment naïve individuals, but very low in individuals who had failures of therapy, significantly lower than in individuals with successful therapy, although the time periods since HIV-1 diagnosis were highest for individuals in group FT, intermediate for those in group NF and shortest for those in the NT group. However, a trend to higher cross neutralization in individuals infected for a shorter period of time appeared to be more relevant than the actual viral loads determined in the year before sample collection, as suggested by the data from the plasma in the study by Kim et al. (2001). Studies have shown that the CD4 T lymphocyte nadir is very important for reconstitution of cellular immune response during/ after HAART (Siddique et al. 2006). An analysis of nadir CD4 levels, however, showed no correlation between these values and the capacity for neutralizing the HIV-1 isolates included in this study. A surprisingly high proportion of individuals with no detectable viral loads for the last year before sample collection showed a good humoral immune response, both in antibodies capable of binding V3 peptides and in antibodies effective in controlling heterologous HIV-1 replication in vitro. Although seroreversion appears to be rare even after years of successful antiviral therapy, the gradual decrease of specific anti-HIV-1 antibodies is a well known occurrence in HAART patients (Béniguel et al. 2004, Amor et al. 2006, Cornelissen et al. 2006). Several hypotheses have been raised to explain the maintenance of a humoral anti-HIV-1 immune response after years of apparent absence of HIV-1 antigenic stimulation, including maintenance of HIV-1 antigens associated to follicular dendritic cells in patients with undetectable viremia (Popovic et al. 2005). Based on the results presented here, therapy failure, similar to rebounds in viremia, will lead to a stimulation of anti-HIV-1 antibodies, able to recognize specific HIV-1 epitopes but apparently unable to effectively control HIV-1 replication, as HIV-1 neutralization does not appear to be significantly stimulated by the increase in antigen availability. However, the lack of autologous neutralization analyses of this study prevents meaningful conclusions upon the effectiveness or clinical benefit for the individual patients. Although no broader neutralization response was seen in individuals of the FT group, it seems possible that years of HAART, while not completely inhibiting viral replication, may at least in part control the highly infectious and "fit" viral subpopulation, and, possibly, give rise to "antigenic debris". These debris (Parren et al. 1997) may be responsible for the induction of a specific immune response observed by binding to synthetic peptides, but are unable to induce production of antibodies capable of interfering with cellular infection by HIV-1. ACKNOWLEDGEMENTS To the patients for participating in this study, to the donors and the Haematology Service of the Clementino Fraga Filho Hospital, UFRJ, for donation of surplus buffy coats. To Dr Eva-Maria Fenyoe for donation of the HIV-1 IIIB reference isolate. The technical assistance of Luciano S Amaral, Vanessa Martins and Giselle Furtado Borges is gratefully acknowledged. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07148f1.jpg] [oc07148t1.jpg] [oc07148f3.jpg] [oc07148f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}