 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 102, No. 8, December, 2007, pp. 931-936 Resistance to experimental autoimmune encephalomyelitis development in Lewis rats from a conventional animal facility Sofia Fernanda Gonçalves Zorzella, Juliana Seger, Douglas Rodrigues Martins, Ana Claudia Pelizon, Alexandrina Sartori/+ Instituto
de Biociências, Departamento de Microbiologia e Imunologia,
Universidade Estadual Paulista, Distrito de Rubião Júnior
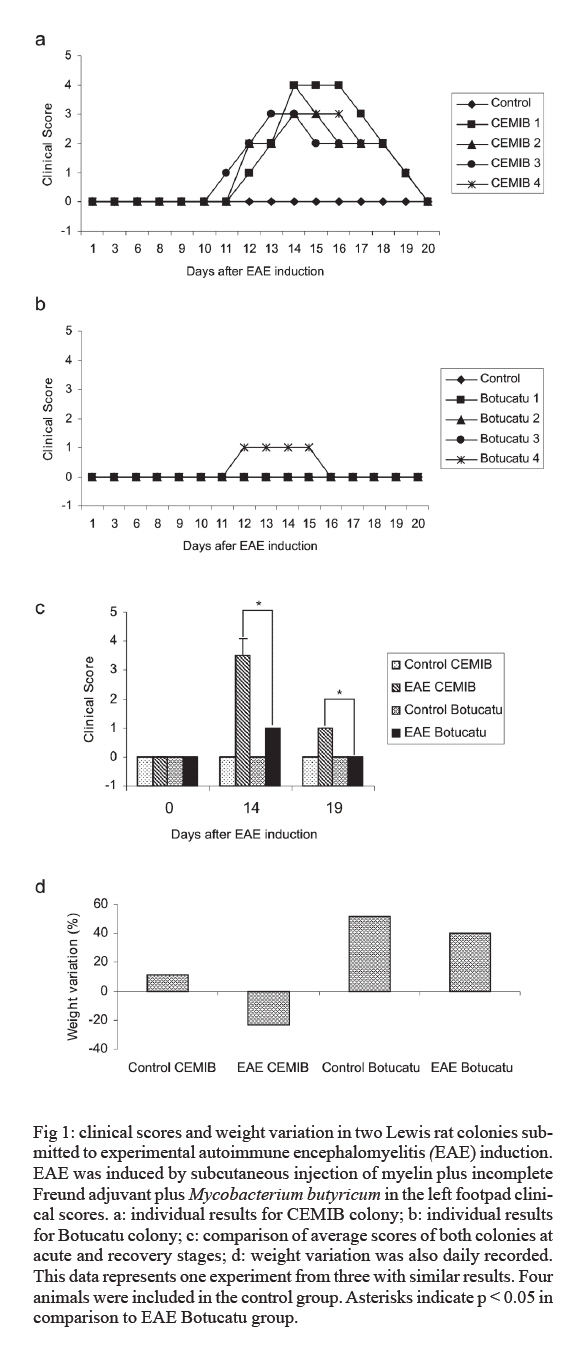
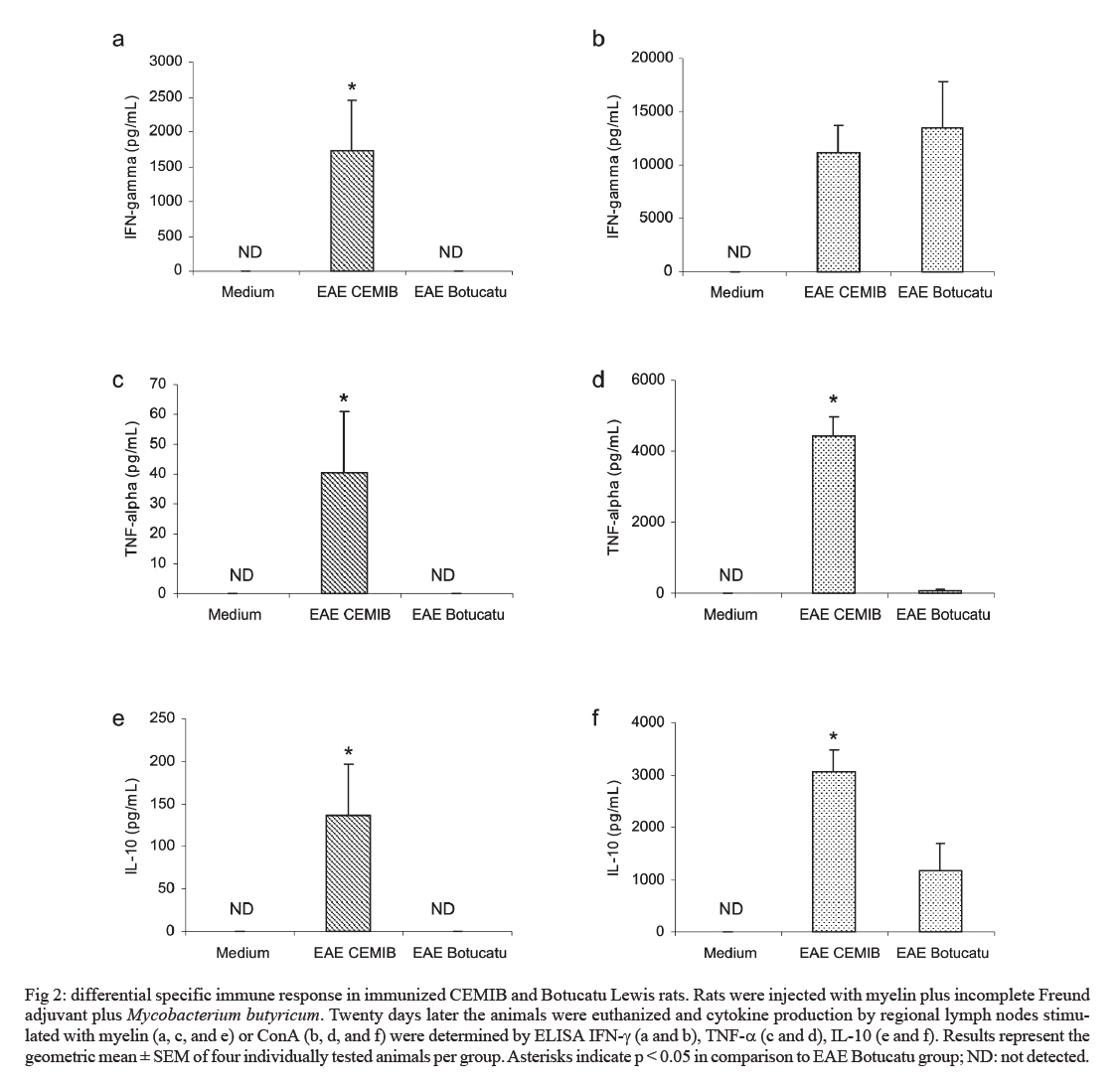
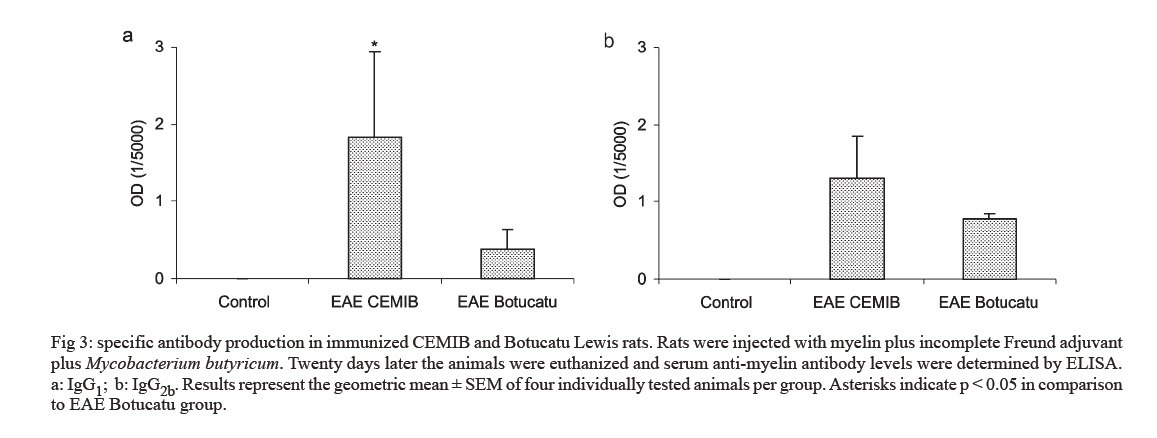
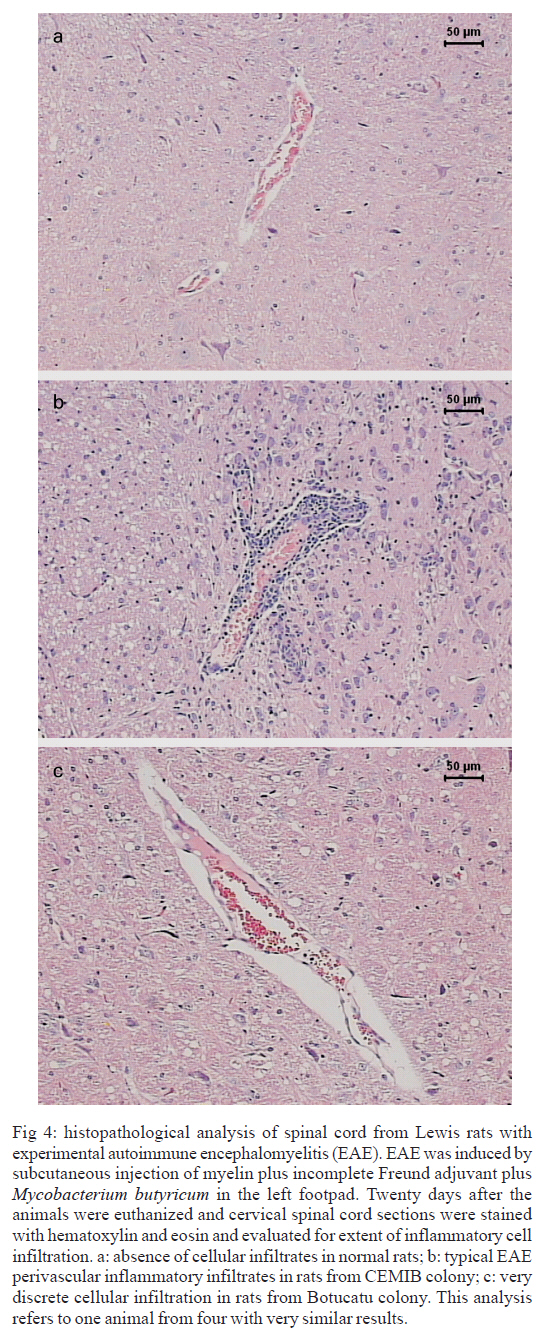
s/n, 18618-000 Botucatu, SP, Brasil Financial support: CNPq, FAPESP. Received 3 July 2007 Code Number: oc07168 Experimental autoimmune encephalomyelitis (EAE) is an inflammatory disease of the brain and spinal cord that is mediated by CD4+ T lymphocytes specific to myelin components. In this study we compared development of EAE in Lewis rats from two colonies, one kept in pathogen-free conditions (CEMIB colony) and the other (Botucatu colony) kept in a conventional animal facility. Female Lewis rats were immunized with 100 μl of an emulsion containing 50 μg of myelin, associated with incomplete Freund’s adjuvant plus Mycobacterium butyricum. Animals were daily evaluated for clinical score and weight. CEMIB colony presented high EAE incidence with clinical scores that varied from three to four along with significant weight losses. A variable disease incidence was observed in the Botucatu colony with clinical scores not higher than one and no weight loss. Immunological and histopathological characteristics were also compared after 20 days of immunization. Significant amounts of IFN-γ , TNF-α and IL-10 were induced by myelin in cultures from CEMIB animals but not from the Botucatu colony. Significantly higher levels of anti-myelin IgG1 were detected in the CEMIB colony. Clear histopathological differences were also found. Cervical spinal cord sections from CEMIB animals showed typical perivascular inflammatory foci whereas samples from the Botucatu colony showed a scanty inflammatory infiltration. Helminths were found in animals from Botucatu colony but not, as expected, in the CEMIB pathogen-free animals. As the animals maintained in a conventional animal facility developed a very discrete clinical, and histopathological EAE in comparison to the rats kept in pathogen-free conditions, we believe that environmental factors such as intestinal parasites could underlie this resistance to EAE development, supporting the applicability of the hygiene hypothesis to EAE. Key words: experimental autoimmune encephalomyelitis - rats inbred Lewis - cytokines The extraordinary susceptibility of the Lewis rat to a large number of different inducible tissue-specific autoimmune diseases has made it a much-studied experimental model in the field of autoimmunity. Experimental autoimmune encephalomyelitis (EAE) is a Th1 mediated inflammatory paralytic disease of the central nervous system (CNS), which serves as a model of human demyelinating diseases of the CNS such as multiple sclerosis (MS) (Swanborg 1975, Gold et al. 2006). The autoreactive CD4+ T cells cross the blood-brain barrier and secrete the pro-inflammatory cytokines IFN-γ and TNF-α in response to epitopes present in CNS proteins, including myelin basic protein (MBP), proteolipid protein and myelin oligodendrocyte glycoprotein. EAE can be actively induced in the Lewis rat by subcutaneous injection of MBP in Complete Freund's Adjuvant (CFA), or passively by the adoptive transfer of in vitro MBP-restimulated T cells from MBP-sensitized syngeneic donor rats (Swanborg 1988). The clinical course of MBP-induced EAE in the Lewis rat is an acute ascending paralysis, that is monophasic and self-limiting, making it an ideal model to study the regulatory events that lead to recovery from autoimmune diseases (Swanborg 1995). MS and EAE development are clearly affected by different factors as gender (Papenfuss et al. 2004), age (Ditamo et al. 2005), and also by genetic and environmental conditions (Sotgiu et al. 2004). The hygiene hypothesis has been proposed to explain the increased incidence of autoimmune diseases and allergy in areas of the world with improved health care and sanitation. This hypothesis proposes that the lack of intense infections in industrialized countries owing to improved hygiene, vaccination and use of antibiotics alters the immune system in a way that it responds inappropriately. Imbalance of Th1 and Th2 responses and lack of regulatory T-cell populations are two of many proposed potential mechanisms for immune failures such as autoimmunity and allergy (Sewell et al. 2002). During standardization of EAE we initially used a Lewis rat colony established in the Department of Microbiology and Immunology (UNESP, Botucatu). In spite of exhaustive trials, only very discrete clinical scores were observed. As our animal facility is conventional and Lewis rats are described as highly susceptible to EAE development (Swanborg 2001), we attributed this resistance to differences in sanitary barrier conditions. To test this possibility we compared EAE development in two colonies of Lewis rats, one raised and kept in pathogen-free conditions (CEMIB colony) and the other in a conventional animal facility (Botucatu colony). MATERIALS AND METHODS Animals - Female Lewis rats (4-6 weeks old) were obtained from two sources: CEMIB/UNICAMP and UNESP/Botucatu, and named CEMIB and Botucatu colonies, respectively. These two colonies are from different origins. The Botucatu colony was established with rats from the laboratory of Clinical and Experimental Allergy and Immunology, Faculty of Medicine, São Paulo University, originally obtained from MA Bioproducts (Walkerville, MD, USA), while the CEMIB colony was established with pairs purchased from Harlan Sprague Dowley (Indianapolis, IN, USA). The animals were fed with a standard granulated food and water ad libitum. Food and water were sterilized. Animals were manipulated in compliance with the ethical guidelines adopted by Colégio Brasileiro de Experimentação Animal, being the experimental protocol approved by the local Ethics Committee (protocol 458). Induction and scoring of EAEThe main immunization protocol used for both colonies was inoculation of 100 µl of an emulsion containing 50 µg of MBP (Sigma Chemical Co, St. Louis, MO, USA) associated with 50 μl of Incomplete Freund's Adjuvant (Difco Laboratories, Detroit, MI, USA) plus 5 mg/ml of Mycobacterium butyricum (Difco Laboratories, Detroit, MI, USA), in the left footpad. Larger amounts of myelin or Freund's adjuvant or subcutaneous route were also tried. Non immunized animals from both colonies were used as control groups. Weight variation and clinical scores were daily recorded. Signs of disease were graded as 0 (zero): no disease; 1: loss of tonicity of the distal portion at the tail; 2: total loss of tail tonicity; 3: hind limb weakness (partial paralysis); 4: complete hind limb paralysis and urinary incontinence, and 5: moribund. Histopathological analysis - The histological analysis of cervical spinal cord sections was performed on day 20 following immunization. Fragments from the cervical spinal cord were removed, fixed in 10% formalin and embedded in paraffin. Five micrometer sections were cut and stained with hematoxylin and eosin and examined with an Olympus microscope. Anti-myelin antibody levels - Serum samples were collected on day 20 following immunization and tested by enzyme linked immunosorbent assay (ELISA) for the presence of antibodies against MBP. Briefly, plates were coated with 5 µg/ml of antigen in coating solution (Na2CO3/NaHCO3, pH 9,6) overnight at 4ºC. Non-specific antibody binding was blocked by incubation with 0,05% Tween 20, 10% fetal calf serum (FCS) in phosphate buffered saline (PBS, 200 µl per well) for 1 h at 37ºC. Subsequently the plates were incubated overnight at 4ºC with dilutions of serum from control and immunized rats. For the detection of specific serum IgG1 and IgG2b, the plates were incubated with biotinylated mouse anti-rat antibodies (Oxford Biotechnology). Plates were then incubated for 30 min at room temperature with Strept AB (kit from Dako, Carpinteria), and revealed by adding H2O2 with o-phenylenediamine (OPD) (Sigma, St. Louis, MO, USA). Color development was stopped with H2SO4 and optical density was measured at 492 nm. IFN-γ, TNF-α and IL-10 production - Normal and immunized rats were sacrificed 20 days after immunization. Lymph node (popliteal + inguinal) cells were collected, and adjusted to 2.5 x 106/ml. Cells were cultured in complete RPMI medium (RPMI supplemented with 5% FCS, 20 mM glutamine and 40 IU/ml of gentamicin), in the presence of 10 µg/ml of myelin or 5 µg/ml of Concanavalin A (ConA, Sigma, St. Louis, MO, USA). Cytokine levels in culture supernatants were evaluated 48 h later by ELISA according to manufacturer instructions (R&D Systems, Minneapolis, MN, USA). 96 well plates (NUNC) were coated with capture antibodies for IFN-γ (DY 585), TNF-α (DY 510) or IL-10 (DY 522) diluted in PBS at 2 µg/ml, 4 µg/ml and 4 µg/ml, respectively. Plates were incubated overnight and then blocked during 2h with 1% albumin in PBS. Standard rat cytokines and culture supernatants were added and the plates incubated for 2 h. Biotinylated anti- IFN-γ , anti-TNF-α and anti-IL-10 antibodies were added (150, 100 and 100 ng/ml, respectively), and plates were incubated during 2 h at room temperature. Then, plates were incubated at room temperature for 30 min with Streptavidin and then revealed by adding H2O2 + OPD. Color development was stopped with H2SO4, and optical density was measured at 492 nm. Sensitivity of ELISA for IFN-γ, TNF-α and IL-10 were 20, 30 and 30 pg/ml, respectively. Statistical analysis - Statistical analysis was performed using Minitab Version 1996 (Minitab, State College, PA, USA). One-way ANOVA and the Tukey test were used to compare the clinical scores, the production of anti-myelin antibodies and the cytokine levels (p < 0.05). RESULTS EAE susceptibility in CEMIB and Botucatu colonies - Susceptibility to EAE development was clearly different in these two colonies. All animals from CEMIB colony developed a standard paralysis, reaching score 4 during the acute phase of the disease (Fig. 1a). As expected from other reports these animals lost around 25% of their initial weight. Differing from these results, animals from Botucatu colony displayed a very low susceptibility to EAE development. In the experiment showed at Fig. 1b, the animals reached only score 1 (paralysis at the tail tip). In addition, these animals manifested no weight loss, and they even showed a weight gain similar to that observed in the control group. Weight variation in both colonies is documented at Fig. 1d. A diversity of protocols, including the use of increased amounts of myelin or M. butyricum, inoculation by subcutaneous route at the dorsal region and immunization of younger animals were additionally tried. Also, some immunized rats from Botucatu colony were observed during prolonged periods. These strategies were incapable of enhancing EAE development in Lewis rats from this colony. Differential immune response to myelin in CEMIB and Botucatu colonies - Lymph node cells from immunized CEMIB rats stimulated in vitro with myelin produced significantly higher levels of IFN-γ, TNF-α and IL-10 (Figs 2a,c, and e, respectively) than cultures from Botucatu colony. IFN-γ production by ConA stimulated cells was similar in both colonies (Figs 2b), whereas TNF-α and IL-10 induction by ConA were also clearly higher in CEMIB colony (Figs 2d and f respectively). Levels of IgG1 (3a), but not of IgG2b (Fig. 3b) were significantly higher in CEMIB colony. Histological analysis - Inflammatory infiltrates were observed in the CNS of both colonies but were quantitatively very distinct. Confirming the clinical status observed in CEMIB colony, a clear perivascular mononuclear cell inflammation was observed in the cervical spinal cords of these animals. In contrast, the corresponding neurological tissues from Botucatu colony showed very discrete inflammatory foci. These differences can be observed in Fig. 4a (normal rat), Fig. 4b (CEMIB EAE rat), and Fig. 4c (Botucatu EAE rat). DISCUSSION A very clear difference in susceptibility to EAE development was observed between the two colonies. CEMIB animals colony developed a standard disease, clinically characterized by significant weight loss and high clinical score during the acute phase of the disease. This high susceptibility has frequently been reported by many authors since the early description of EAE in Lewis rats (Swanborg 1975, Stepaniak et al. 1995, Gold et al. 2006). These clinical manifestations were also associated with typical immunological and histopathological signs of EAE development. IFN-γ and TNF-α levels whose activity is considered pivotal to both, MS and EAE evolution (Mustafa et al. 1991, Imitola et al. 2005), were significantly elevated in lymph node cell cultures stimulated with myelin. The severity of histologic neural tissue lesions clearly correlated with these clinical symptoms. Cervical spinal cord sections showed typical and abundant inflammatory foci, localized around small vessels, similar to many research reports (Stanislaus et al. 2001). Differing from these results, animals from Botucatu colony immunized in similar conditions, displayed a very low susceptibility to EAE development. These animals presented no weight loss and even showed a weight gain similar to the one observed in the control group. They also showed very low clinical scores characterized by loss of tonicity only at the distal portion of the tail. Other protocols of EAE induction, including the addition of higher amounts of autoantigen, adjuvant and the use of a subcutaneous route of immunization did not change the low clinical scores observed in Botucatu colony. Analysis of the immunological parameters clearly demonstrated that specific anti-myelin response was much more prominent in the CEMIB colony. The finding of a significantly higher production of IFN-γ and TNF-α in these animals that developed EAE with high scores is supported by many literature reports that attribute a fundamental role of these cytokines to CNS lesions in both, EAE and MS (Mustafa et al. 1991, Imitola et al. 2005). High IL-10 production by the CEMIB colony was expected and is supported by reports that demonstrate its presence during spontaneous clinical recovery (Diab et al. 1997), and also during immunomodulatory procedures that decrease clinical and histopathological alterations (Stanislaus et al. 2002). However, differently form what was expected, animals form Botucatu colony submitted to EAE induction produced no IL-10 in response to myelin and even produced less IL-10 after ConA stimulation. We cannot, however, exclude other possibilities as production of this cytokine by other kind of cells as dendritic cells or by cells that migrated to the CNS as has been demonstrated in EAE (Jander et al. 1998, Yang et al. 2000). Additionally, the contribution of other cytokines as TGF-β that is described as an important endogenous mechanism to limit the extension of inflammation cannot be ruled out (Tanuma et al. 1997). In this context, our data could suggest that disease resistance may be a result of an immunologic deficit, maybe triggered by a regulatory T cell (Treg cell). Contribution of Treg cells to initiate remission phase in experimental EAE and its deficiency in clinical MS manifestations have been reported (Viglietta et al. 2004, Mann et al. 2007). Therefore, a very appealing explanation for this striking difference could be the differential sanitation condition used to raise and keep these two colonies. This possibility is based on a growing body of evidence suggesting that excessive hygiene conditions could contribute to the increased incidence of allergic and autoimmune diseases in developed countries. This idea has been named " hygiene hypothesis " and has been supported by investigations in epidemiology (Cooke et al. 2004), basic immunology (Vercelli 2006), animal models (Kitagaki et al. 2006), and human trials with probiotic agents (Floch & Monrrose 2005). A prominent theme emerging from this research has been the discovery of a relative deficiency of T-regulatory cell activity in allergy and autoimmunity (Randolph & Fathman 2006). Additionally, in some animal models, restitution of T-regulatory cell activity has been achieved by exposure to infectious agents such as helminths or products derived from them (Wilson et al. 2005). The relevance of hygiene hypothesis to EAE and MS has been scientifically supported by both experimental approaches and epidemiological studies in MS population. For example, in a murine EAE model, it was demonstrated that the infection with Schistosoma mansoni delayed the onset of the disease and also prevented inflammation in the CNS (La Flamme et al. 2003, 2004, Sewell et al. 2003). Epidemiological investigation revealed a dichotomous relationship between the global distribution of MS and parasitic infections (Fleming & Cook 2006). Also, longitudinal and migratory studies of MS have been consistent with the hygiene hypothesis, showing a paralleled increase in MS and improved sanitation (Cabre et al. 2005). In this context we evaluated the presence of worm eggs in the feces and adult worms in the intestine of Lewis rats from both colonies. Interestingly, a high amount of eggs from Aspiculuris sp. and Syphacia sp. were found in Botucatu animals, but not in CEMIB ones. In light of recent reports that demonstrated or suggested a protective effect of Schistosoma infections in MS or EAE, these results support the view that this natural rodent infection could protect these animals from developing EAE. The possibility that a helmintic infection can block EAE development in Lewis rats from CEMIB colony is being investigated in our laboratory. This approach will allow us to establish the contribution of the infection and also of Treg cells to EAE resistance as has been demonstrated for allergic airway inflammation (Wilson et al. 2005). Even though we believe that our findings and the epidemiological evidences suggest a strong correlation with the hygiene hypothesis, we cannot exclude the possibility of a genetic diversion in these animals. Genetic mutations involving, for example, nitric oxide synthase and regulatory cytokines, which have been described as relevant factors for EAE development, could contribute to this resistance (Cautain et al. 2001, Staykova et al. 2005). REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07168f2.jpg] [oc07168f4.jpg] [oc07168f3.jpg] [oc07168f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}