 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
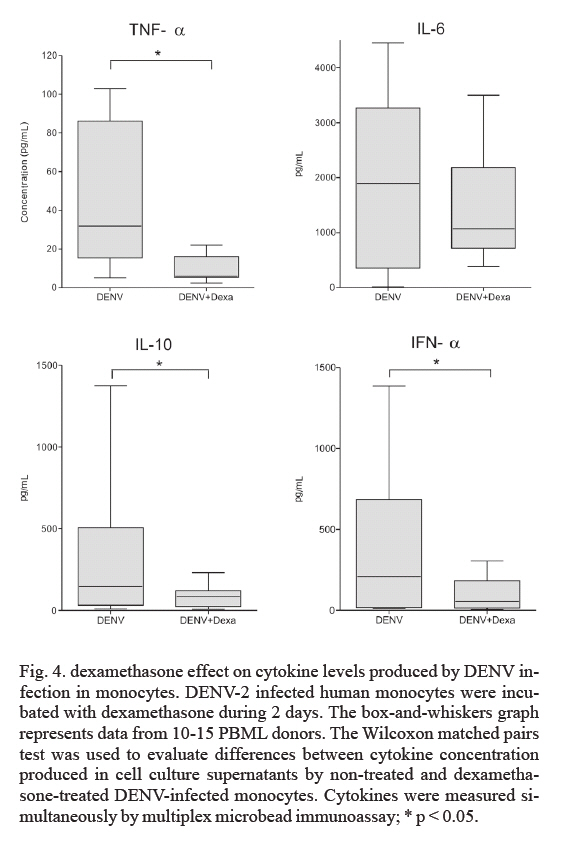
Memórias do Instituto Oswaldo Cruz, Vol. 102, No. 8, December, 2007, pp. 983-990 An in vitro model for dengue virus infection that exhibits human monocyte infection, multiple cytokine production and dexamethasone immunomodulation Sônia Regina Nogueira Ignácio Reis, André Luiz Franco Sampaio*, Maria das Graças Muller Henriques*, Mariana Gandini, Elzinandes Leal Azeredo, Claire Fernandes Kubelka/+ Laboratório
de Imunologia Viral, Instituto Oswaldo Cruz-Fiocruz *Departamento
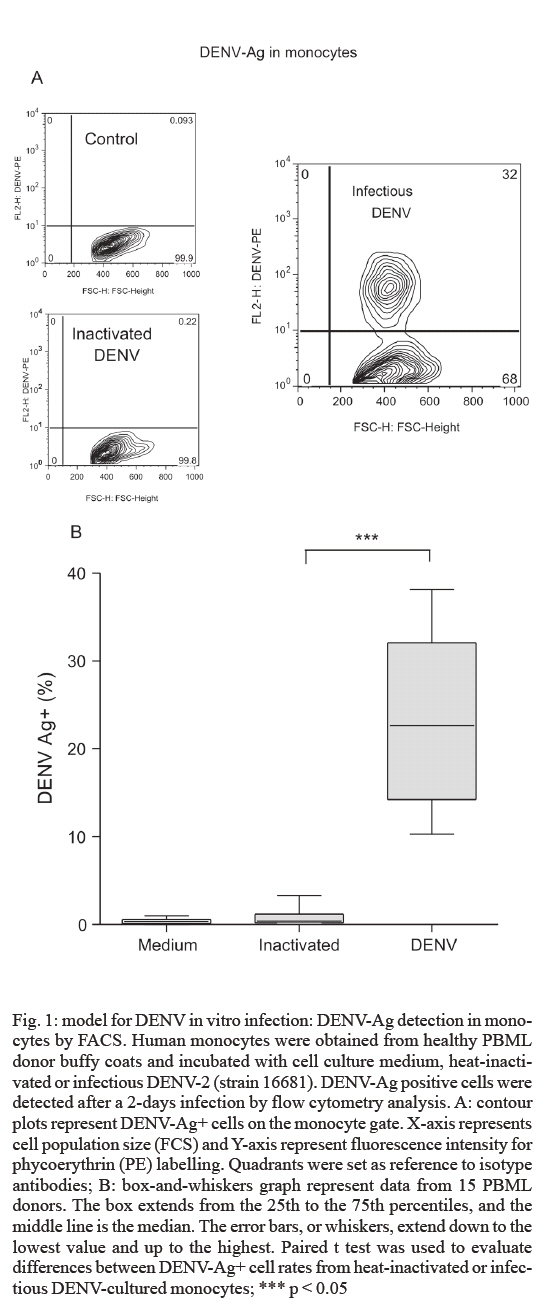
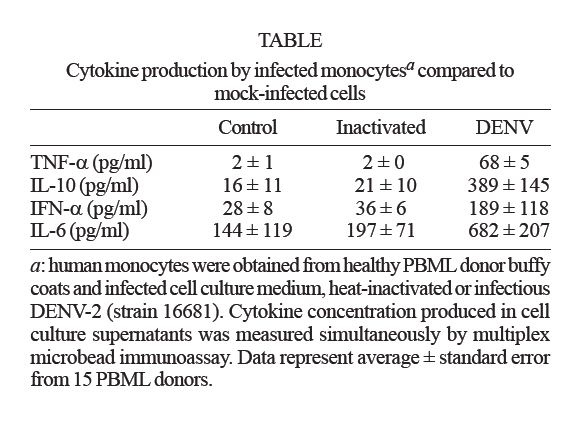
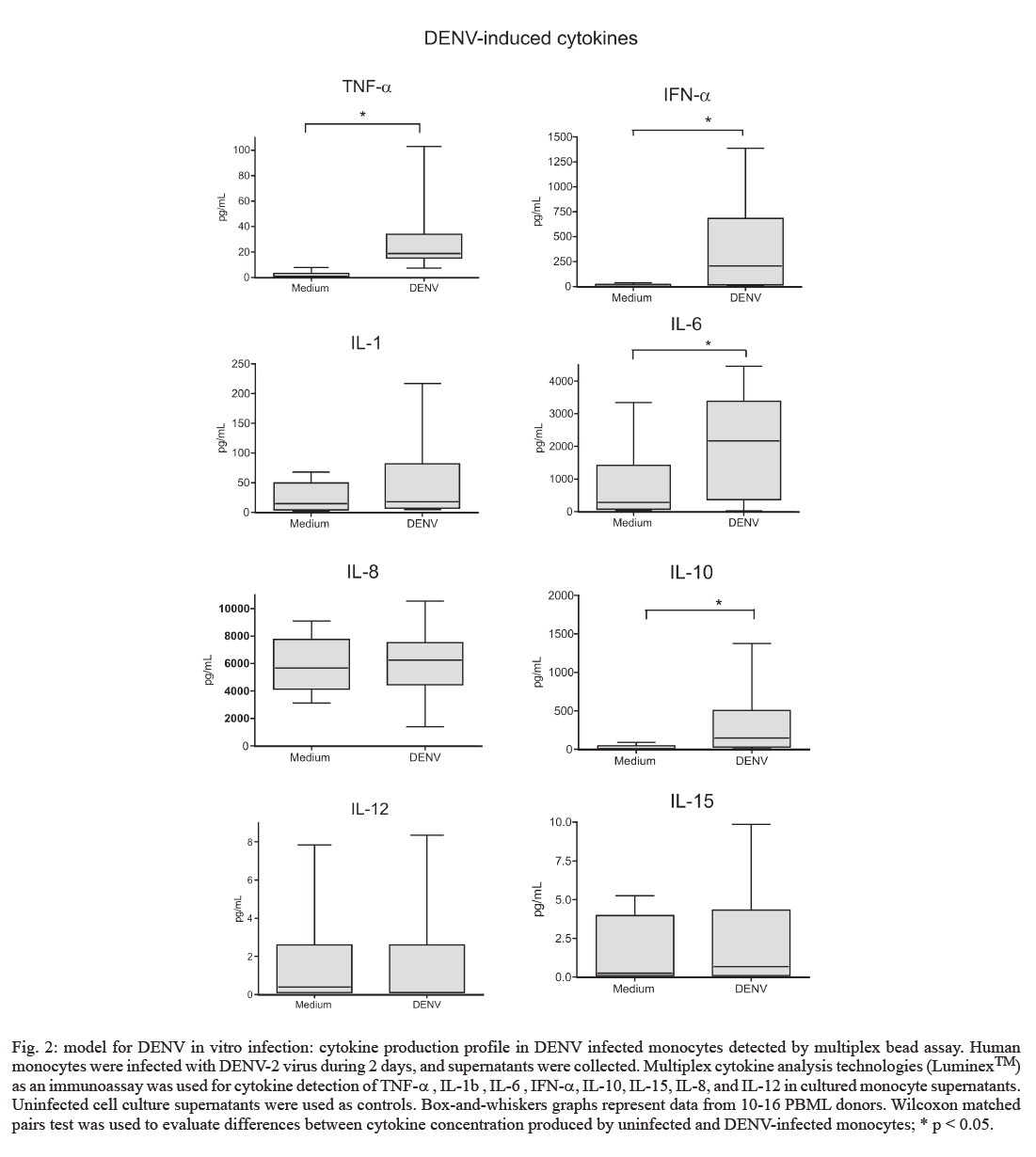
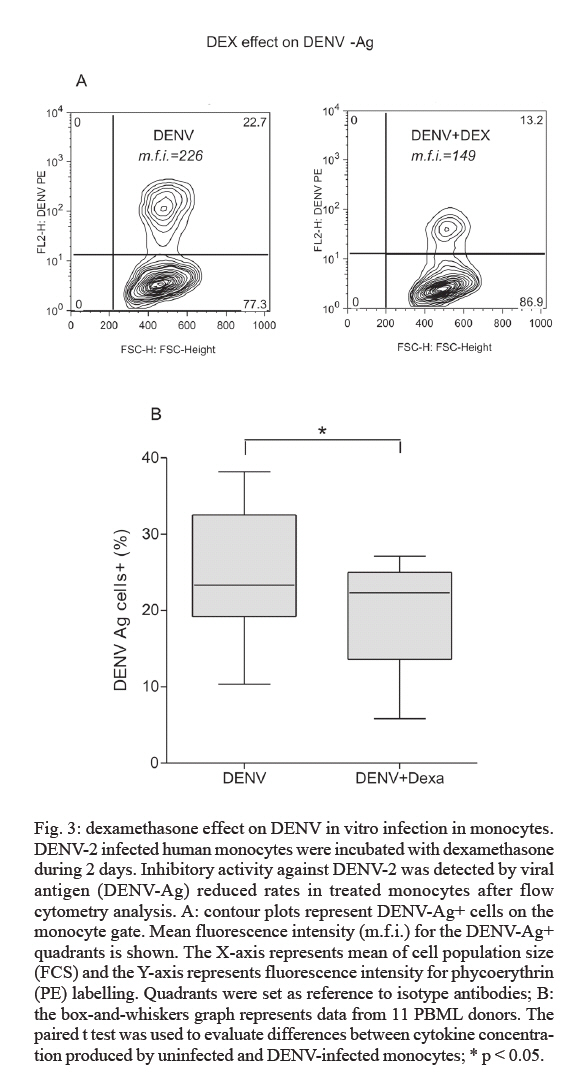
de Farmacologia Aplicada, Financial support: PDTIS-Fiocruz, CNPq (50.1567/03-8), ICGEB (CRP.LA/ARG03-01) Received 20 August 2007 Code Number: oc07175 An important cytokine role in dengue fever pathogenesis has been described. These molecules can be associated with haemorrhagic manifestations, coagulation disorders, hypotension and shock, all symptoms implicated in vascular permeability and disease worsening conditions. Several immunological diseases have been treated by cytokine modulation and dexamethasone is utilized clinically to treat pathologies with inflammatory and autoimmune ethiologies. We established an in vitro model with human monocytes infected by dengue virus-2 for evaluating immunomodulatory and antiviral activities of potential pharmaceutical products. Flow cytometry analysis demonstrated significant dengue antigen detection in target cells two days after infection. TNF-a, IFN-a, IL-6 and IL-10 are produced by in vitro infected monocytes and are significantly detected in cell culture supernatants by multiplex microbead immunoassay. Dexamethasone action was tested for the first time for its modulation in dengue infection, presenting optimistic results in both decreasing cell infection rates and inhibiting TNF-a, IFN-a and IL-10 production. This model is proposed for novel drug trials yet to be applyed for dengue fever. Key words: dengue - dexamethasone - monocytes - cytokines - therapeutics Dengue fever (DF) is an acute infectious viral disease that presents a broad severity spectrum from asymptomatic or oligosymptomatic to extremely severe clinical manifestations. It is believed that immunological mechanisms play a key role in pathogenesis (WHO 1997, Gubler 2002). Proinflammatory cytokines have been associated with haemodynamic and coagulation disorders that may lead to increased vascular permeability and leakage and to hypovolemic shock resulting eventually in death (Green & Rothman 2006). No specific treatment exists for dengue and research has been unsuccessful in finding improved conditions with steroids, antivirals or substances that decrease capillary permeability (Sumarmo et al. 1982, Tassniyom et al. 1993, Pea et al. 2001, Gibbons & Vaughn 2002, Ligon 2005). In patients, fluid-replacement therapy should be administrated according to the severity and paracetamol/metamizole has been prescribed for fever and analgesia. Glucocorticoids are widely adopted as anti-inflammatory drugs for several inflammatory diseases and are known to suppress of inflammatory mediators expression (Barnes & Adcock 1995, Joyce et al. 1997). Dexamethasone inhibits cytokine gene expression such as TNF-α by blocking transcription factors NF-kB and activator factor-1 (Steer et al. 2000). IL-1β , iNOS, ciclooxigenase-2 and monocyte chemoattractant protein-1 have also been described to destabilize mRNAs during this treatment (Amano et al. 1993, Ristimaki et al. 1996, Poon et al. 1999, Lasa et al. 2001, Korhonen et al. 2002). Dexamethasone is applied in the clinics to treat pathologies with inflammatory and autoimmune origins, such as rheumatoid arthritis, multiple melanomas and bacterial meningitis (Jimenez-Zepeda & Dominguez-Martinez 2006, van de Beek & de Gans 2006, Schutt et al. 2007) despite some described adverse effects (Stoll et al. 1999, Hempen et al. 2002, Vardy et al. 2006). Reports concerning corticosteroids are yet inconclusive regardin their benefits during DF (Panpanich et al. 2006). Herein we present an in vitro model with human primary monocytes infected with dengue virus serotype 2 (DENV-2). We evaluated virus load by detecting antigen positive cell rates through flow cytometry and cytokine secretion profile after cell infection by a multiple fluorescent microbead immunoarray. This model is proposed for DF novel drug trials, and dexamethasone was tested for the first time, presenting successful modulation by decreasing infected monocyte frequencies and inhibiting TNF-α , IFN-α and IL-10 production in cell culture supernatants. MATERIALS AND METHODS Cell cultures, preparation of virus stock and virus titration - An Aedes albopictus C6/36 cell clone was grown as monolayers at 28ºC on Leibovitz medium (L-15) supplemented with 200 mM glutamine, 1% non-essential amino acids solution, 0.5% tryptose phosphate broth, 100 U/ml penicillin, 10 mg/ml streptomycin and 5% foetal calf serum (FCS). DENV-2 strain 16681 was provided by Dr. Scott B. Halstead (Naval Medical Research Center, USA). The virus was titrated by serial dilution cultures in microtiter plates and detected by immunofluorescence as previously described (Miagostovich et al. 1993). Virus titre was calculated as 50 percent tissue culture infectious dose (TCID50/ml) (Reed & Muench 1938, Schoepp & Beaty 1984). The inactivated virus was prepared by incubating the inoculum for 30 min/56ºC. Titrated virus stock was obtained at a concentration of 5.5 x 1010 TCID50/ml. Preparation of human peripheral blood mononuclear leukocytes (PBMLs) - Human peripheral blood was obtained from the Hospital Universitário Clementino Fraga, Rio de Janeiro. Human PBMLs from healthy donors were isolated from buffy coat cells through density gradient centrifugation (350 g/30 min in Ficoll-Paque Plus, Amersham Biosciences Corp) according to standard procedures. Cells were suspended in RPMI 1640 supplemented with 200 mM glutamine, 100 U/ml penicillin and 10 mg/ml streptomycin, afterwards incubated at 37ºC under humid atmosphere with 5% CO2 and allowed to adhere on uncoated polystyrene flasks (150 cm3) during 90 min for monocyte enrichment. Non-adherent cells were removed by washing, and adherent cells were detached by mechanical cell harvesting with cell scrapers in cold cell culture medium, then resuspended in supplemented RPMI plus 10% FBS. Infection of adherent PBMLs and treatment with dexamethasone - Enriched monocytes suspended in supplemented RPMI 1640 medium plus 10% FCS were seeded at 2 x 106 cells/ml on 96 or 24-well plates. After an overnight incubation, infection was performed with inoculum diluted in cell culture medium (30 or 300 ml) containing 2.2 x 109 TCID50/ml. After a 2 h-incubation period for adsorption, the cell culture supernatant was replaced with cell culture medium plus 2% FCS and incubated with dexamethasone 0,05 mM at 37ºC with 5% CO2. After 48 h, supernatants were collected and stocked at -20ºC until cytokine measurement and cells were recovered for viral antigen determination by flow cytometry. Cell viability was verified in culture by Trypan blue exclusion and presented > 95% viability. Wells with cell control, inactivated and infectious DENV were assayed. Cytokine detection in cell culture supernatant by multiplex microbead immunoassay - A multiplex biometric immunoassay, containing fluorescent dyed microspheres conjugated with a monoclonal antibody specific for a target protein, was adopted for cytokine measurement according to the manufacturer's instructions (Upstate), measured cytokines being: IL-1β , IL-6, CXCL8 (IL-8), IL-10, IL- 12 (p70), IL-15, IFN-α, and TNF-α. Briefly, cell culture supernatant (50 ml) was incubated overnight with antibody-coupled beads. Complexes were washed and then incubated with biotinylated detection antibody and streptavidin-phycoerythrin prior to assessing cytokine concentration titres, the concentrated human recombinant cytokine provided by the vendor. A broad range 1.95 - 8,000 pg/ml recombinant cytokines was used to establish standard curves as well as maximize assay sensitivity and dynamic range. Cytokine levels were determined using a multiplex array reader from Luminex™ Instrumentation System (Bio-Plex Workstation from Bio-Rad Laboratories). The analyte concentration, calculated with the software from the manufacturer (Bio-Plex Manager Software), provided a regression analysis to derive the equation prediction of the cytokine concentrations in plasma samples. Determination of viral antigen by flow cytometry in monocytes - Adherent monocytes were recovered by scraping with a plastic microtip using cold cell culture medium, then set at 1 x 106 cells/microtube, centrifuged (350 g/10 min) and washed once with 1ml phosphate-buffered saline pH 7.4 containing 1% bovine serum albumin with 0.1% NaN3 (PBS/BSA). Cells were fixed with a solution containing 2% paraformaldehyde PBS/BSA at 4ºC/20 min and permeabilized with a solution containing 0.15% saponin in PBS/BSA. Permeabilized cells were then blocked with 5% inactivated plasma in PBS/BSA at 4ºC/30 min and incubated with mouse anti-dengue complex monoclonal antibody (Chemicon, USA) at 4ºC/60 min. Cells were then washed and incubated with anti-mouse IgG labelled with phycoerythrin (DAKO, USA) for 30 min/4ºC. After incubation, cells were washed with PBS/BSA and resuspended in 2% paraformaldehyde. Cells were acquired (5,000 events for gated monocytes) on a FACS® Calibur flow cytometer (Beckon & Dickinson, USA) and analysed with FlowJo Software (TreeStar Inc, CA, USA). Isotype-matched antibody was adopted as a staining negative control. Statistical analyses - Data were firstly tested for normality with the GraphPad Prism version 4.02 for Windows, GraphPad™ Software (San Diego, CA, USA, http://www.graphpad.com) in order to determine the significance of differences in DENV antigen positive (DENV-Ag+) cell rates under culture treatment conditions. Data values passed the Kolmogorow-Smirnov normality test and were evaluated for significance using the paired t test (two-tailed). However, the cytokine detection data values obtained by multiplex bead assay did not pass the normality test and was therefore analysed by the non-parametric Wilcoxon matched pairs test (two-tailed). Altered parameters were considered significant at p < 0.05. RESULTS Human monocytes as an in vitro infection model for DENV: infected cell rates and cytokine production profile - Human monocyte-enriched PBMLs were infected with DENV-2 (strain 16681) and cultured in vitro. In preliminary experiments we searched for DENV-Ag+ cells by flow cytometry analysis (FACS), which indicated that our DENV stock (tested at 2.2 x 109 TCID50/ml) induced infection rates peaking on the second day after infection using three different PBML donors (data not shown). Target cells are CD14+ cells (data not shown), which constitute approximately 95% cells in the monocyte region from cell population size and granularity (FCS x SSC) dotplots in FACS analysis as described previously (Neves-Souza et al. 2005). We also observed that only infectious DENV, but not heat-inactivated DENV, was effective in DENV-Ag detection (Fig. 1A). Heat-inactivated DENV incubation with monocytes presented data equivalent to those of cell culture medium indicating that the antigen detected when the infectious virus is present might be a result of virus replication. Infected monocytes from 15 different PBML donors varied, however DENV-Ag expression was statistically significant when compared with heat-inactivated DENV (Fig. 1B). Infection rates from the second day ranged from 10 to 38% total monocytes. After DENV monocyte infection, cell culture supernatant was collected and tested for the presence of eight different cytokines by multiplex microbead immunofluoroscent assay (LuminexTM technology), while the mock inoculum with heat-inactivated DENV did not stimulate monocytes cytokine production (Table). In 10-16 PBML donors tested on the second day after infection, TNF-α, IL-6, IL-10 and IFN-α were detected at statistically significant, increased levels when compared to control monocyte supernatants (Fig. 2). These cytokines were still present in subsequent days varying their detection levels, while other cytokines (IL-1β, IL-8, IL-12, and IL-15) did not alter their levels during seven days in tested cultures. The two-day time point was determined to evaluate drug effect for both antiviral and immunomodu-lating activities. Dexamethasone-induced reduction of DENV-Ag+ cell rates on monocytes detected by FACS - Dexamethasone has been reported to be an immunomodulating agent and experimentally demonstrated to decrease viral titres, as well as reducing virus-induced inflammation (Moreno et al. 2003). DENV infected human monocytes from 11 different PBML donors were treated with dexamethasone for two days in culture. DENV-Ag+ cell expression was detected by FACS (Fig. 3A) and exhibited a highly significant inhibiting effect on infected cell frequencies (Fig. 3B). Hence, the dexamethasone effect results in viral protein downregulation, indicating that virus particle load is decreased in monocytes after treatment. Dexamethasone-induced inhibition of cytokine detection in DENV infected monocyte supernatants - Glucocorticoids, such as dexamethasone, inhibit the expression of inflammatory mediators such as pro-inflammatory cytokines (Joyce et al. 1997). The dexamethasone immunomodulating activity was tested on DENV-infected human monocytes cultured for two days. TNF-α, IL-6, IL-10, and IFN-α were measured in cell supernatants from 10-15 different PBML donors with the multiplex bead immunoassay. Significant TNF-α, IL-10 and IFN-α inhibition was achieved in treated cultures in contrast to untreated (Fig. 4). IL-6 levels in supernatants from dexamethasone-treated cell cultures were not statistically different from infected control cultures, although eight out of 15 PBML donors presented > 35% IL-6 inhibition rates (total averages: untreated, 1,883 ± 379 and dexamethasone-treated, 1,347 ± 244 pg/ml), five had unchanged levels and two had > 35% increased levels. Dexamethasone was acknowledged to be an effective in vitro treatment to downregulate cytokines produced during monocyte infection by DENV. DISCUSSION Clinical investigations report an important role for cytokines during DF pathogenesis. Among others, TNF-α, IL-1β, IL-6 and IL-8 have been associated with severity in various studies (Vitarana et al. 1991, Hober et al. 1993, Bethell et al. 1998, Raghupathy et al. 1998, Braga et al. 2001, Fink et al. 2006). Cytokines can be related to haemorrhagic manifestations (Azeredo et al. 2001), coagulation activation, fibrinolysis (Suharti et al. 2002) and may be implicated in vascular permeability and worsening morbidness (Green & Rothman 2006). These conditions are most striking in severe patients, nevertheless may appear during mild disease as well (Avila-Aguero et al. 2004, Nguyen et al. 2004). Several target cells have been designated by different investigators to perform DENV infection studies in vitro. Mononuclear phagocytes, such as dendritic cells and monocytes, have been appointed as relevant cells for infection both in vitro and in vivo (Scott et al. 1980, Hotta et al. 1984, Halstead 1988, Wu et al. 2000). Circulating peripheral monocytes from patients during the acute phase present DENV-Ags indicating their role during natural infection in DF (Neves-Souza et al. 2005). Here we present an in vitro model that detects DENV positive cell frequencies in human primary monocytes and four different cytokines, which are participating in disease development. We demonstrated that after in vitro intervention with dexamethasone this model is successfully modulated for DENV-Ag expression and cytokine production. TNF-α (Espina et al. 2003) and IL-8 (Bosch et al. 2002) production by human primary monocytes were earlier reported after DENV infection. In vitro differentiated macrophages secrete TNF-α , IFN-α, IL-1β, IL-8, IL-12, MIP-1α, and RANTES after infection but not IL-6, IL-15, or nitric oxide (Chen & Wang 2002). We infected freshly isolated human monocytes from 15 healthy PBML Brazilian donors, and our results revealed that DENV induced in these target cells a different cytokine pattern, TNF-α, IFN-α, IL-6, and IL-10 being upregulated as detected earlier in DF patients. On the other hand IL-1β, IL-8, IL-12, and IL-15 were not significantly detected in monocyte cultures, these cytokines being produced at low levels during DF in most Brazilian patients studied (unpublished observations) and remarkable absent in reports (Green & Rothman 2006) except for IL-8. IFN-a is markedly elevated during defervescense in dengue haemorrhagic fever (DHF) (Kurane et al. 1993). Since the DENV-2 strain used is the same as that from Chen and Wang's report, we can speculate that differences in cytokine production may be related to target cell origin and that monocytes may change their cytokine production after in vitro differentiation. In addition, cytokine genetic polymorphism may also explain these differences considering that the former study was performed in Southeast Asia, in contrast to our South American population, which corroborates the evidence that DF profiles change among several geographic/ethnic regions (Moraes et al. 2006). Some authors (Chareonsirisuthigul et al. 2007) have exhibited the simultaneous production of several cytokines after DENV infection as herein delineated, but their target cells were derived from a transformed cell line, not primary isolated monocytes. We describe for the first time the dexamethasone inhibitory effects for both DENV-induced cytokine production and virus load reduction on monocytes. There have been several reports of in vitro DENV infection inhibition by different treatments, but human monocytes or other primary cells were not described as targets in these earlier studies. Some treatments inhibited the glycan-processing enzyme endoplasmic-reticular glycosidase and the morphogenesis of viruses that bud from the endoplasmic reticulum (Gu et al. 2007). Others, such as the virus inhibitor ribavirin, act on the enzyme IMP dehydrogenase (IMPDH) that catalyzes the de novo biosynthesis of guanine nucleotides affecting dengue replication (Markland et al. 2000). It is still unknown whether or not the antiviral effect observed here is a direct effect on virus replication steps or an indirect effect stimulating antiviral cytokines/factors. We demonstrated a novel inhibitory effect induced by dexamethasone that has a striking impact on TNF-α, IFN-α and IL-10 production in human monocyte cultures after DENV-2 infection. This drug is known for its modulating activity on TNF-α production and NF-kB nuclear translocation, which is an essential step for TNF-a induction on stimulated monocytes (Steer et al. 2000). We suggest that NF-kB may exert as well an activation function on TNF-α synthesis after DENV infection and may be one target for dexamethasone. IL-1β (Jeon et al. 2000), IL-6 (Amano et al. 1993), and IL-10 (Bessler et al. 2001) are inhibited by dexamethasone after monocyte induction by LPS, a bacterial product involved in septic shock. Contrastingly, IL-6 is induced during DENV in vitro infection, but dexamethasone does not change significantly its levels in culture. This lack of effect on IL-6 by dexamethasone may be undermined due to the evidence that no important role was found for IL-6 in Brazilian patient severity during recent epidemics (unpublished observations). Other cytokines were also associated with severity, such as IL-18 and TGF-β (Azeredo et al. 2006), and the dexamethasone effect deserves to be studied for these factors as well. We observed IFN-a downregulation, which may represent an effective immunomodulatory procedure by dexamethasone. This effect is not interfering with virus clearance which can be explained by the antiviral effect from other molecules such as nitric oxide, known to control DENV replication in monocytes (Neves-Souza et al. 2005, Chareonsirisuthigul et al. 2007). IL-10 is known for its blocking effect on STAT-1 and IRF-1 (Mahalingam & Lidbury 2002), relevant factors for iNOS activation (Dell'Albani et al. 2001), and its expression during DENV infection may result in enzyme inhibition together with reduced nitric oxide effect on DENV replication. After IL-10 downregulation by dexamethasone, iNOS activation may occur, generating nitric oxide that would more efficiently exert its antiviral effects on monocytes. We can not rule out that the antiviral effect was induced by the drug direct interference on virus replication or that cytokine downregulation may be a result from less intense monocyte activation by the virus. Further investigation on theses mechanisms deserve to be performed. One must be aware of adverse effects described in clinical immunotherapeutic procedures during some severe infectious diseases (Hempen et al. 2002, Vardy et al. 2006). Although dexamethasone presented very interesting effects on the in vitro dengue infection model described here, there is insufficient clinical evidence to justify the use of corticosteroids in managing dengue shock syndrome. A recent report evaluates death, blood transfusion need, convulsions and pulmonary haemorrhage concluding that there was no overall benefit attributed to corticosteroids, but the number of participants in the analysis was small (Panpanich et al. 2006). We believe that this model will be useful to track new substances with therapeutic properties, however further investigations will be mandatory in order to achieve a safe immunomodulatory intervention in which pro-inflammatory factors will be downregulated to a moderate level still enabling virus replication control. This would result in more efficient restoration of host homeostasis, ultimately consummating in avoidance of the most deleterious clinical manifestations during DF. ACKNOWLEDGEMENTS To Dr. Jussara P. Nascimento (In memoriam) for her constant encouragement. To the technical support of Alessandro Souza, Maryrose Lavatori, and Mariana Lopes. To Mitchell Raymond Lishon for reviewing the manuscript. REFERENCES
Copyright 2007 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc07175f4.jpg] [oc07175t1.jpg] [oc07175f3.jpg] [oc07175f2.jpg] [oc07175f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}