 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 1, February, 2008, pp. 39-44 Patterns of co-association of C-reactive protein and nitric oxide in malaria in endemic areas of Iran Hossein Nahrevanian/+, Jafar Gholizadeh1, Mahin Farahmand, Mehdi Assmar Department
of Parasitology, Pasteur Institute of Iran, Pasteur Avenue, 13164
Tehran, Iran 1Urumia Laboratory of Education and Quality
Control, Health Center of West Azerbaijan Province, Urumia, Iran Financial support: Pasteur Institute of Iran (project n. 200) Received
12 June 2007 Code Number: oc08006 In addition to numerous immune factors, C-reactive protein (CRP) and nitric oxide (NO) are believed to be molecules of malaria immunopathology. The objective of this study was to detect CRP and NO inductions by agglutination latex test and Griess microassay respectively in both control and malaria groups from endemic areas of Iran, including Southeastern (SE) (Sistan & Balouchestan, Hormozgan, Kerman) and Northwestern (NW) provinces (Ardabil). The results indicated that CRP and NO are produced in all malaria endemic areas of Iran. In addition, more CRP and NO positive cases were observed amongst malaria patients in comparison with those in control group. A variable co-association of CRP/NO production were detected between control and malaria groups, which depended upon the malaria endemic areas and the type of plasmodia infection. The percentage of CRP/NO positive cases was observed to be lower in NW compare to SE region, which may be due to the different type of plasmodium in the NW (Plasmodium vivax) with SE area (P. vivax, Plasmodium falciparum, mixed infection). The fluctuations in CRP/NO induction may be consistent with genetic background of patients. Although, CRP/NO may play important role in malaria, their actual function and interaction in clinical forms of disease remains unclear. Key words: C-reactive protein - Iran - nitric oxide - malaria - Plasmodium
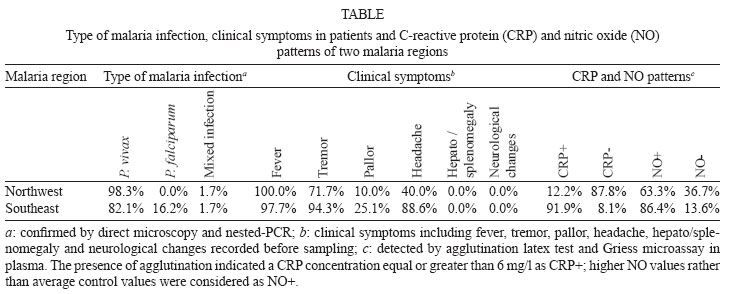
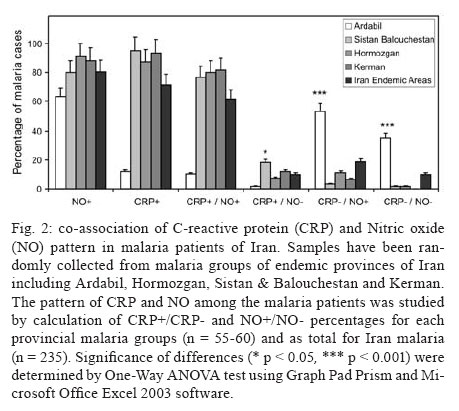
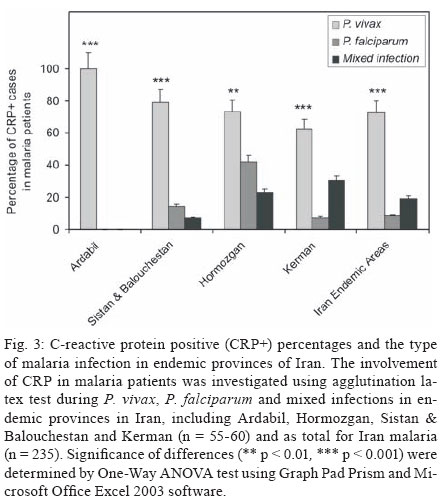
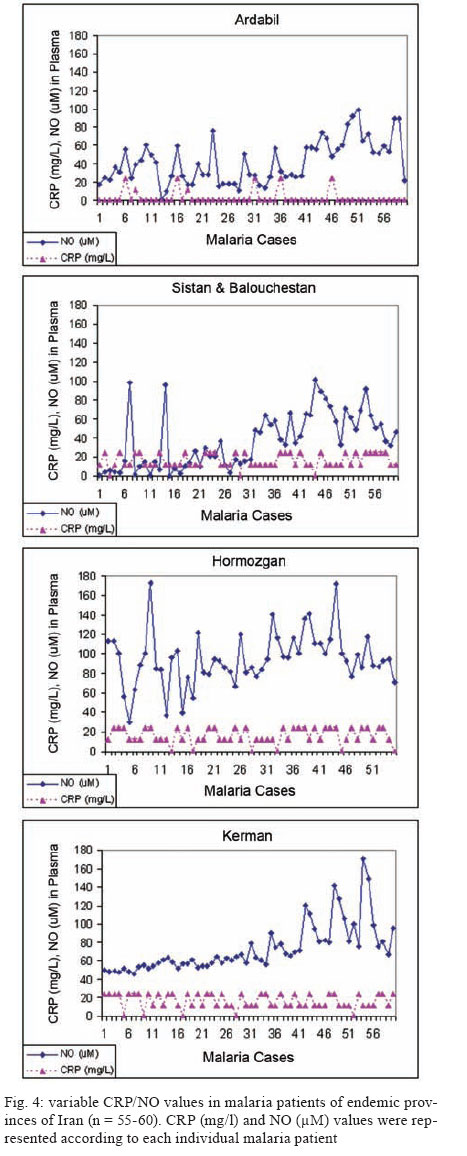
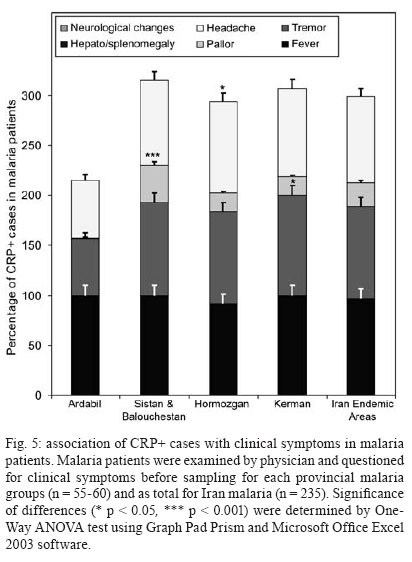
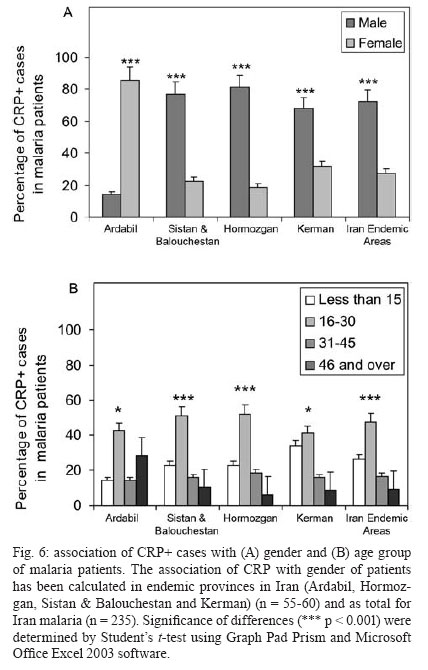
Malaria is a world wide problem and it still remains as a concern in the Eastern Mediterranean region and in Iran (Sadrizadeh 1999, WHO 2003). Iran was divided into two malaria zones, North and South of the Zagros Range of Mountains (Manouchehri et al. 1992), including Southeastern (SE) and Northwestern (NW) provinces (Zakeri et al. 2004, Nahrevanian et al. 2006). The prevalent species is Plasmodium vivax followed by Plasmodium falciparum (Zaim 1987, Edrissian 2002) and mixed-infections (Assmar et al. 2003). Out of 23,562 malaria cases have been reported in the country in 2003, more than 70% occurred in the SE part (Zakeri et al. 2002, WHO 2003). Recently, a new threat of imported malaria emerged from the NW part of the country, Parsabad area, which was affected by a serious epidemic of P. vivax (Sadrizadeh 1999). To date, the malaria situation in SE corner is serious (Zakeri et al. 2002). However, in other endemic areas, malaria is hypoendemic with uncomplicated cases (Edrissian et al. 1993). A malaria eradication programme began in Iran in 1949 and changed to malaria control in 1985 as a result of constraints and challenges. In the SE provinces, the major peak of malaria transmission occurs between September and November, with 21% of malaria cases in this region caused by P. falciparum (WHO 2004). Chloroquine (CQ) is still being recommended as the first-line treatment for uncomplicated cases of P. falciparum malaria, with a combination of sulfadoxine-pyrimethamine and quinine recommended for the CQ treatment failures. A combination of CQ and primaquine is the recommended first-line treatment for P. vivax malaria (Raeisi et al. 2006). Vector control activities include residual house spraying, larviciding, biological control and insecticide-treated bed nets distribution (WHO 2004). Increased drug and insecticide resistance has made vaccine preparation urgent for malaria with a focus on new immune targets. Potential mechanisms of immunity against malaria include antibodies, cytotoxic T-cells, cytokines and a variety of other soluble mediators. Macrophages are key components of the antimicrobial immune responses, generating large amounts of toxic molecules, reactive oxygen and nitrogen intermediates (RNI), H2O2and nitric oxide (NO) (Bogdan et al. 2000). In addition to NO, C-reactive protein (CRP) is a major acute phase protein present in normal serum, which increases significantly after most forms of tissue injuries and infections as a non-specific innate defense mechanism of the host. CRP as a protein is mainly regulated at the transcriptional level induced by cytokines (Kremsner et al. 1996, Ablij & Meinders 2002). It is a marker of inflammatory reactions and cytokine activation (Jakobsen et al. 1998), which is produced very early after infection (McCarty et al. 2004). Besides CRP being a marker of the inflammatory process, it may also play a modulating role at the site of inflammation by their effect on the expression of different adhesion molecules. The CRP levels have been increased several times after the acute event with a strong positive correlation between the duration and the intensity of the stimulus and the number of hepatocytes synthesizing CRP (Ablij & Meinders 2002). CRP is reported to be a critical element during malaria infection and there is a strong association of elevated CRP levels during the acute phase of severe (Kremsner et al. 1996) and clinical forms of malaria (Jakobsen et al. 1998). Although, the role of NO, RNI and NO synthase (NOS) were investigated previously in rodent malaria (Nahrevanian & Dascombe 2001, 2002, Dascombe & Nahrevanian 2003) and human malaria (Nahrevanian et al. 2006), the co-association between NO and other immune factors in clinical forms of malaria remains controversial (Balmer et al. 2000, Cramer et al. 2004). The importance of a balance in the chemokine network was outlined for a protective immunity against malaria infection. The balance includes the amounts of cytokine released, the rate, time and site of production. Interaction, intervention and co-association throughout this network are both interesting and complicated. In addition to NO induction which was reported previously in Iranian malaria patients (Nahrevanian et al. 2006), in this novel immunoepidemiological study, the pattern of co-association between CRP and NO is investigated in malaria patients and control groups from NW and SE endemic malaria regions of Iran. PATIENTS, MATERIALS AND METHODS Patients and groups - This study has been carried out from 2002 to 2005 to investigate CRP and NO patterns among the malaria patients and control groups from Hormozgan, Sistan & Balouchestan, Kerman (SE) and Ardabil (NW) as malaria endemic provinces (Fig. 1). Two hundreds and thirty five blood samples (55-60/province) were randomly collected from malaria patients referred to the health clinics, who resided in above mentioned endemic areas. In addition, 80 blood samples (20/province) were randomly taken from healthy people with no history of malaria from same endemic regions. Sample selection in both groups was applied considering all statistical parameters including province, gender (control group = male 51%, female 49%; malaria group = male 53%, female 47%) and age groups (control; 30.3 ± 1.9 and malaria; 28.4 ± 1.5). Malaria was confirmed by microscopy and polymerase chain reaction assay (PCR), and all clinical symptoms including fever, tremor, pallor, headache, hepato-splenomegaly and neurological changes were recorded (Table). This study protocol was approved by institutional ethical review board (Ethical Committee of the Pasteur Institute of Iran). Blood collection and plasma preparation - 10 ml of peripheral venous blood was collected from malaria patients in two sets of tubes (with EDTA for DNA extraction and with Heparin for CRP and NO assays) as described previously (Nahrevanian et al. 2006). Malaria diagnosis - Malaria was diagnosed by both microscopy (Giemsa stained thick and thin blood smears) and PCR in patients as previously described (Nahrevanian et al. 2006). Up to 100 microscopic fields were examined to establish the diagnosis. Plasmodia DNA was extracted from blood samples by nested-PCR as described by Snounou et al. (1993). First round of PCR was carried out using rPLU5 and rPLU6 primers. PCR products were reacted with second round primers including FAL1, FAL2 (P. falciparum), VIV1, VIV2 (P. vivax) and MAL1, MAL2 (Plasmodium malariae) (Snounou et al. 1993). DNA was amplified according to the following condition: denaturing 95ºC/1 min, annealing 58ºC/2 min and extension 72ºC/2 min for 35 cycles. PCR products were run in a 1.5% agarose gel, and observed by UV-transluminator. Agglutination latex test (ALT) for CRP assay -ALT was based on slide agglutination and applied as recommended by kit manufacturer (SGM, Roma, Italy). Positive and negative control were placed into separate circles on the slide test. The CRP latex reagent was gently swirled and added and mixed to the tested blood sample, and spread over the entire surface of the circle by a mechanical rotator. Latex particles with goat IgG anti-human CRP were agglutinated when mixed with samples containing CRP. The presence or absence of visible agglutination was examined macroscopically and the presence of agglutination indicated a CRP concentration equal or greater than 6 mg/l. Griess microassay (GMA) - A modified Griess reaction was used (Rockett et al. 1994 modified by Nahrevanian & Dascombe 2001). Standard curves for sodium nitrite and nitrate (Sigma, UK) were prepared. Samples were treated with nitrate reductase (NAD[P]H Aspergillus species, Sigma, UK) and NADPH β-nicotinamideadenine dinucleotide phosphate (Sigma Diagnostics, St Louis, USA). Griess reagent [5% phosphoric acid, 1% sulfanilic acid and 0.1% N (1-naphthyl-1)-ethylendiamine dihydrochloride, all from Sigma, UK, dissolved in 100 ml deionised water] was then added and proteins subsequently precipitated by trichloroacetic acid (BDH, England). Tube contents were mixed then centrifuged (Eppendorf centrifuge 5415 C, Germany) and duplicates of supernatants were transferred to a flat-bottomed microplate, and absorbances read at 520 nm using a microplate reader (Bio-TEK, power wave XS, USA) and values were calculated from standard calibration plots (Nahrevanian & Dascombe 2001). Statistical analysis - Values are presented as the mean ± SEM for groups of n samples. The significance of differences was determined by Analysis of Variances (ANOVA) and Student's t-test using Graph Pad Prism Software (Graph Pad, San Diego, California, USA) and Microsoft Office Excel 2003. RESULTS The results indicated that CRP and NO are produced in all malaria endemic areas of Iran. In this study, NO level of patients was compared with that of the related controls in each province, whereas higher NO values rather than average control values were considered as NO positive (NO+) (Nahrevanian et al. 2006). In addition, the presence of agglutination indicated a CRP concentration equal or greater than 6 mg/l as CRP positive (CRP+). Both data from GMA and ALT were calculated and established for a cross comparison amongst study groups. Data revealed more CRP/NO positive cases among malaria patients in comparison to healthy controls. In addition, a higher rate of CRP+ (91.9%) and NO+ (86.4%) cases was observed in SE than NW malaria [CRP+, 12.2%; NO+, 63.3%]. It indicated an association between patterns of CRP+/NO+ cases with geographical distribution of malaria in Iran. This may be related to the different type of malaria in the NW (mostly P. vivax with a minority of mixed infections) and SE regions (P. vivax, P. falciparum and mixed infections) (Table). Despite of similar patterns of CRP/NO involvement among SE provinces, a different result was found in NW province (p < 0.001). In spite of similar patterns of CRP/NO involvement among SE provinces, a different result was presented in NW province (p < 0.001). A consistent association was observed among all CRP/NO positive and CRP/NO negative cases (Fig. 2). In addition, a relationship was observed between CRP+ cases and the type of malaria infection. The highest association of CRP was observed in P. vivax malaria as a prevalent species, then it was presented in mixed infections of both plasmodia; whereas the lowest involvement was indicated in P. falciparum malaria (p < 0.01, p < 0.001) (Fig. 3). A different variation was detected in CRP and NO values between two control and malaria groups. Variable patterns for CRP (mg/l) and NO (µM) production were represented according to each individual of malaria patients in four endemic provinces. The fluctuations in CRP/NO values induction by different population may be consistent with genetic differences of human profiles (Fig. 4). The results demonstrated an association between CRP+ cases and clinical symptoms mostly fever, tremor and headache, however some variations were observed in provinces including tremor (p < 0.05), pallor (p < 0.001) and headache (p < 0.05) (Fig. 5). A reverse association of CRP positive cases with gender was observed between NW malaria (male 14.3%; female 85.7%: p < 0.001) and SE malaria (male 75.4%; female 24.6%: p < 0.001) regions (Fig. 6A). According to age groups, the CRP+ cases in 1-15 years of malaria patients were 14.3% (NW) and 26.5% (SE); in 16-30 years were 42.8% (NW) and 48.0% (SE); in 31-45 years were 14.3% (NW) and 16.9% (SE) and in over 45 years old were 28.6% (NW) and 8.6% (SE). The major CRP induction was associated with 16-30 years age group in all endemic regions (p < 0.001) (Fig. 6B). DISCUSSION In this novel immuno-epidemiological study, CRP and NO levels were higher in malaria groups, when compared with controls of same endemic area. The detected high CRP level in malaria in this study is in agreement with previous reports, and it is hence proposed as a useful tool for malaria immuno-epidemiology. The resulting data has revealed a correlation of CRP and NO in malaria patients of NW and SE Iran. Variations arise due to the geographical areas and the type of malaria infection. This may clarify the co-involvement of CRP and NO as two major immune elements during malaria infection in endemic regions of Iran; however it is not justified, whether the CRP/NO production is beneficial or detrimental to the patients. Kremsner et al. (1996) reported the high plasma levels of both CRP and NO in patients with severe malaria than uncomplicated cases. Moreover, stimulated hepatocytes produce CRP, which has an anti-plasmodial effect on the hepatic development of parasite, both by preventing penetration of the sporozoite and by blocking parasite replication (Nussler et al. 1991). Notwithstanding the conflicting publications, the role of CRP (Kremsner et al. 1996, Gyan et al. 2002) and NO (Awasthi et al. 2003, Clark et al. 2003, Cramer et al. 2004) in the immune responses to Plasmodium remains uncertain. It is suggested that NO alone or in accompany with CRP and/or other chemokines is involved in protective and/or pathogenic responses of human malaria. However, there are substantial data in the literature on CRP and NO suggesting a potent antimicrobial role for both of them individually or acting together. Increased CRP/NO synthesis might have a protective rather than pathological role in malaria (Gillman et al. 2004, Sharma et al. 2004). Moreover, some authors suggested no association with degree of disease and induction of CRP/NO in malaria. These findings did not support a pivotal role for systemic generation of CRP/NO in the pathogenesis of severe malaria (Clark et al. 2003). Some researchers believe that NO (Becker et al. 2004) and CRP or their toxic functions may contribute to the pathology of severe form of disease, and that many of malaria symptoms can be depended on excessive CRP induction (Kremsner et al. 1996, Jakobsen et al. 1998) or NO overproduction (Gyan et al. 2002, Clark et al. 2003). CRP was associated with the pathological signs including fever, splenomegaly, anaemia and cerebral malaria (Gyan et al. 2002). In addition, the duration of symptoms and parasitemia correlated positively with CRP during the acute phase of malaria (Kremsner et al. 1996). Data of this study emphasized that CRP/NO involvement in malaria varied and depended on endemic regions and strain of Plasmodia or many other unknown factors. Variation of genetic structure and polymorphisms of genes encoding CRP/NO induction could describe the ability of host responses to malaria infection, which are reported in different publications of various regions (Ikeda et al. 2002, Clark et al. 2003, Wells et al. 2005). The variations in CRP/NO induction by different population may be consistent with the human genetic background. The protective polymorphisms may undergo selection in populations with a long history of exposure to malaria and other infections (Boutlis et al. 2003). In addition, population studies generally support an association between protection from severe malaria and CRP/NO production. Genetic epidemiology or cytokine biology alone is not enough to solve CRP/NO paradox, but together they stand a good chance (Gyan et al. 2002, Clark et al. 2003, Mackintosh et al. 2004, Wells et al. 2005). The higher percentage of NO+ /CRP+ cases in SE provinces may indicate a dependency of NO/CRP induction with type of malaria infections (P. falciparum or mixed infection). However further investigation is needed to clarify this concept. In conclusion, our data highlighted the fact that CRP/NO are produced in malaria patients of all endemic provincial areas of Iran. The results of published reports clarified the variation rate of CRP/NO production, which were relating to type of infection, provincial regions, species of Plasmodium and clinical symptoms. The present study was not designed to answer questions relating to the role of CRP/NO in protection or pathogenesis during malaria; our findings provided a frame work of immuno-epidemiological investigations between basal CRP/NO production in two healthy and malaria groups in Iran. Although, CRP/NO play an important role in malaria, their actual functions and interactions in clinical forms of disease remain unclear. However, involvement of CRP/NO in malarial host is conflicting (Boutlis et al. 2004, Wells et al. 2005), the complex relationship between malaria symptoms, genetic polymorphisms and CRP/NO production in populations requires further studies to address their immuno-modulatory roles in malaria. ACKNOWLEDGEMENTS To the Director and the staff from the malaria unit of the Centre for Diseases Control (CDC), Iran, for their cooperation. To the authorities and staff from malaria endemic provinces of Iran during this study. To Dr Vahid Khalaj from Department of Biotechnology, Pasteur Institute of Iran, for reviewing the manuscript and providing helpful suggestions. REFERENCES
Copyright 2008 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08006t1.jpg] [oc08006f4.jpg] [oc08006f2.jpg] [oc08006f1.jpg] [oc08006f6.jpg] [oc08006f3.jpg] [oc08006f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}