 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 1, February, 2008, pp. 98-103 First genotyping of Giardia lamblia from human and animal feces in Argentina, South America Marta C Minvielle, Nora B Molina, Daniela Polverino, Juan A Basualdo/+ Cátedra
de Microbiología y Parasitología, Facultad de Medicina,
Universidad Nacional de La Plata, Calle 60 y 120 s/n, CP 1900, La
Plata, Argentina. Finacial support: Agencia Nacional de Promoción Científica y Tecnológica, Comisión de Investigaciones Científica de la provincia de Buenos Aires, Universidad Nacional de La Plata Received
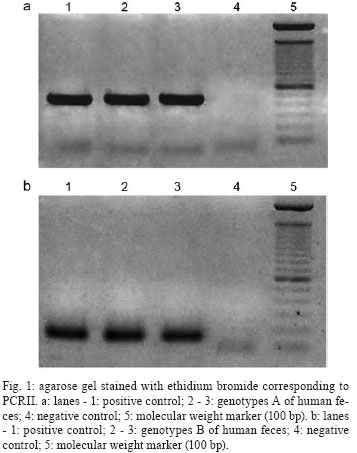
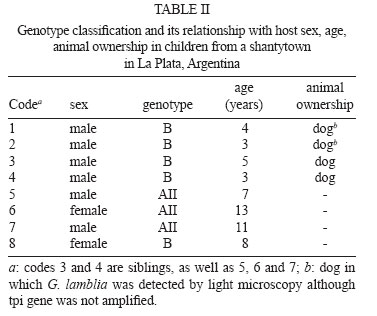
31 October 2007 Code Number: oc08015 The purpose of this study was to investigate the genotypes of Giardia lamblia from human and animal feces and their epidemiological and clinical characteristics in Argentina, South America. Seventy isolates, 60 from humans (adults and children), eight from dogs and two from cows were processed by polymerase chain reaction-restriction fragment length polymorphism. Data corresponding to demographic, socio-cultural and environmental variables and presence/absence of signs/symptoms were collected. The triosephosphate isomerase gene was amplified from 43 (71.66%) of the 60 human fecal samples. Among these, 3/43 (6.98%) were genotype AII and 40/43 (93.02%) were genotype B. Assemblage AII was detected in three children who lived together in a shantytown and they were oligosymptomatic and none had diarrhea. This genotype was not found in animals. Genotype B showed a high prevalence in both adults and children. It was also found in polysymptomatic people, many of whom presented diarrhea. It was also found only in one dog. The present study represents the first contribution to the knowledge of G. lamblia genotypes in Argentina. Key words: Giardia - genotypes - Argentina - South America. Giardia lamblia is a flagellated eukaryotic unicellular microorganism that produces diarrheal disease throughout the world (Adam 2001). This intestinal protozoan has been found in a wide range of mammals and has been accepted as a zoonotic agent (Hopkins et al. 1997, McGlade et al. 2003). In recent decades, molecular classification has been implemented to identify this parasite protozoan and to understand the pathogenesis and host range. Small-subunit rRNA, triosephosphate isomerase (tpi), beta giardin and glutamate-dehydrogenase genomic sequences have been previously studied to mainly permit better identification and comparisons of G. lamblia isolates (Mahbubani et al. 1998, Monis et al. 1999, Adam 2000, Caccio et al. 2002, Lu et al. 2002, Rimhanen-Finne et al. 2002). All of these studies have confirmed the division of human isolates into two major groups called genotype A (or Polish) and genotype B (or Belgian) (Mayrhofer et al. 1995). In addition to their genetic differences, both genotypes can have important number of biological differences. For example, genotype B was significantly more pathogenic in human volunteer infections than genotype A (Adam 2001). Homan and Mank (2001) reported a strong correlation between the mild, intermittent type of diarrhea and assemblage A and severe, actual/persistent diarrhea and assemblage B. However, Eligio-García et al. (2005) and Lalle et al. (2005) did not find any correlation between symptoms and genotype. Genotyping studies have identified G. lamblia isolates from dogs, some of which were found to be genetically identical to those obtained from humans (van Keulen et al. 2002). Studies on G. lamblia obtained from cattle have proved that some isolates belong to genotype E and others to genotype A, and only the latter can cause human infection (O' Handley et al. 2000). About 200 million people in the world are infected with G. lamblia and each individual eliminates up to 900 million cysts per day (Crompton 1999). Higher prevalence is found in tropical and subtropical areas, where G. lamblia affects up to 30% of the population. In epidemiological studies carried out in Argentina, a prevalence of 10% has been found in urban areas and 34% in shantytowns (Gamboa et al. 2003). In rural areas, giardiasis frequency has been close to 4% (Minvielle et al. 2004). In animals from this region the reported prevalence is 7.5% in cattle, 18.6% in dogs, and 3.9% in rodents (Pezzani et al. 2003). Presence of G. lamblia has also been observed in environmental samples (soil and water) from different areas of Argentina (Pierangeli et al. 2003). In Argentina, current methods for the detection of this parasite both in human and animal feces and in environmental samples are usually based on morphological recognition by light microscopy or the use of indirect immunofluorescence with monoclonal antibodies. No previous studies on the use of molecular techniques for G. lamblia in our country were found in the revised bibliography. The aim of this study was to determine the genotype of G. lamblia isolates from human and animal feces (dogs and cows) from La Plata (LP) and General Mansilla (GM), Argentina by using polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) technique and analyzing their clinical and epidemiological characteristics. SUBJECTS, MATERIALS AND METHODS Samples - Fecal samples from 60 (1 to 33-year old) people and ten animals (2 cows and 8 dogs), in which cysts had been detected by light microscopy were analyzed by the modified Telemann technique (Gamboa et al. 2003), including lugol staining. Only one time-related inclusion criterion was considered: samples obtained in population studies (people with and without signs/symptoms) in 2003-2004. All samples were preserved in 5-10% formaldehyde. Participants and legal guardians were requested to give their voluntary consent for the perfonmance of this study. Forty-nine human fecal samples from GM, a rural town 90 km from Buenos Aires, and 11 from a shantytown in LP, 60 km from Buenos Aires, were collected. The fecal samples of six dogs from GM, two dogs from LP and two cows from GM were included. Specific data about risk factors for parasitic infection, including demographic, socio-cultural, and environmental variables, and as well as the presence/absence of signs/symptoms were collected from each human participant. Data corresponding to the following variables were selected: demographic: sex and age; socio-cultural: residence (urban or rural); sanitary condition of the houses (good, medium or poor). "Good" included masonry building, cement/tile floors, fully-equipped bathroom, running water, sewage system, and garbage collection. "Medium" included masonry, cement/tile floors, fully-equipped/half equipped bathroom, home water pump, cesspool and no garbage collection (garbage buried or burnt). "Poor" included wooden/cardboard and/or tin building, dirt floors, latrine, water from a community pump or tap, and open-sky garbage disposal. Overcrowding (yes or no) meant three or more people sleeping in the same room. Environmental: house flooding (never, sometimes, or frequently), where "sometimes" flooding was up to twice a year and "frequently" flooding ocurred three or more times a year. Clinical: presence/absence of signs/symptoms (yes or no), where diarrhea, vomiting, abdominal pain, appetite loss, weakness and sleeping disorders were registered. Parasitic: presence of other parasites in the feces and perianal swab sampling. Cyst purification and disruption - 10 ml of feces filtered through gauze were centrifuged for 5 min at 200 g; 5 ml of phosphate-buffered saline (PBS) were added and centrifuged for 5 min at 200 g. The supernatant was discarded; 3 ml of PBS and 4 ml of sucrose solution (density 1.275) were added. All samples were centrifuged for 5 min at 200 g. The sucrose phase was aspired and transferred to another tube with two volumes of PBS. Cysts were concentrated by centrifugation at 250 g for 5 min. Sediment was resuspended with PBS and stored at 4ºC until use. Cysts were subjected to six cycles of freezing-thawing (-80ºC/+ 8ºC, 30 min) and incubated with Proteinase K (1 mg/ml) and lysis buffer (NaCl 0.15 M, EDTA 0.1 M, SDS 0.5%, pH 7-8) at 37ºC for 24 h (Eligio-García et al. 2002, Polverino et al. 2004). This suspension was stored at -20ºC until DNA isolation. DNA isolation - All buffers and reagents used in this step were provided in the Qiagen® spin-column miniKit (QIAamp DNA stool minikit, Cat. 51504). DNA extraction was performed on 200 ml samples. Two modifications of this protocol were employed: lysis temperature was increased to 95ºC; the AE elution buffer was incubated for 10 min. PCR-RFLP - The tpi gene was amplified according to Amar et al. (2002) (Table I) with modifications: 3mM MgCl2, 0.1 mg/ml bovine serum albumin and 0.1 U/ml Taq DNA polymerase (Invitrogen). All reactions involved an initial denaturation step at 94ºC (4 min), 30 cycles at 94ºC (30 s), 52ºC (30 s) and 72ºC (1 min), with a final elongation step at 72ºC (10 min). A Techgene thermocycler (Techne, Germany) was used. Both positive and negative controls were included in each PCR to validate results (Molina et al. 2007). Positive DNA control genotypes A and B courtesy of Dr. Van Keulen, Cleveland University, and bidistillated water as negative control were used in each PCR. RFLP analysis was performed as described by Amar et al.(2002). Amplification was assessed by electrophoresis of PCR samples on 1.5% or 3.2% agarose gels depending on the expected sizes of amplified DNA. RESULTS The tpi gene was amplified from 43 (71.66%) of the analyzed 60 human fecal samples (Fig. 1a, b). Among humans isolates, 40/43 (93.02%) were genotype B and 3/43 (6.98%) were genotype AII. Genotype classification and its relationship with host sex, age and animal ownership are shown in Tables II and III. Only 1/10 (10%) animal isolates could be genotypified. Genotype AII was detected in three people from LP and genotype B was present in five people from LP and in 35 from GM. All samples with genotype AII were from children (14 years old), while genotype B was present in children as well as adults. Concerning sex, 66.66% of genotype AII and 62.57% of genotype B were found in males. Among children, 92.30% of boys and 92.85% of girls presented genotype B. Three individuals presenting genotype AII lived in urban areas and had poor sanitary conditions, while 87.5% of the people with genotype B lived in rural areas and had either poor (60%-24/40), medium (30%-12/40) or good (10%-4/40) sanitary conditions. Overcrowding was observed in all cases of genotype AII, while only 30% (12/40) had this condition among genotype B. House-flooding was never present among individuals with genotype AII. Individuals with genotype B had frequent flooding in 40% (16/40) of the cases, sometimes in 20% (8/40) and never in 40% (16/40). In order to determine the presence of other parasites, we found that 15 out of 43 people were only infected with G. lamblia (14 from GM and 1 from LP). All genotype-AII infected people were coinfected with Blastocystis hominis. In the case of genotype B, 37.5% (15/40) were coinfected with Enterobius vermicularis, 32.5% (13/40) with B. hominis, 17.5% (7/40) with Entamoeba coli, 10% (4/40) with Ascaris lumbricoides, 5% (2/40) with Trichuris trichiura, and 5% (2/40) with Strongyloides stercoralis. All people with genotype AII reported only weakness and appetite loss. Those with genotype B reported abdominal pain 24/40 (60%), diarrhea 20/40 (50%), appetite loss 19/40 (47.5%), sleeping disorders 15/40 (37.5%), weakness 13/40 (32.5%), and vomiting 10/40 (25%). Among people infected only with G. lamblia (100% genotype B), 9/15 (60%) reported diarrhea, 7/15 (46.66%) abdominal pain, 6/15 (40%) sleeping disorders, 6/15 (40%) weakness, 6/15 (40%) appetite loss, and 5/15 (33.33%) vomiting. Two children (13.33%) reported no signs/symptoms. Among adults studied (all genotype B), one female who had S. stercolaris and G. lamblia, reported sleeping disorders and appetite loss; a male coinfected with B. hominis reported sleeping disorders, abdominal pain and appetite loss; and a female with E. coli, E. vermicularis and B. hominis had no signs/symptoms. DISCUSSION The present study provides, for the first time in our country, information on the distribution of G. lamblia genotypes A and B in both humans and animals from LP and GM. The proportion of samples in which the tpi gene could not be amplified was higher than that reported by Amar et al. (2002), using the same primers. In our study, this could be explained by the presence of PCR inhibitors in some of the fecal samples resulting in prevention of amplification. The failures in the amplification would derive from the low quality DNA of the samples, either due to their degrading in time or because of chemical modifications caused by several substances, formalin among them (Molina et al. 2007). Also, these failures could be the result of the fact that the proportion of DNA in the feces samples was not enough to counteract the effect of the inhibitors that would have co-purified with the nucleic acids (Molina et al. 2007). Surveys in several countries showed a diverse prevalence of genotypes A and B. Studies carried out in Germany, China, Uganda, New Zealand, Egypt, Portugal, and Mexico reported a predominance of genotype A (Karanis & Ey 1998, Yong et al. 2000, Graczyk et al. 2002, Learmonth et al. 2003, Ceu Sousa & Poiares da Silva 2004, El Shazly et al. 2004, Lalle et al. 2005). On the other hand, Read et al. (2004) studied 23 people in Australia and found genotype B in 14, AII in 5, and AI in 4 individuals. In Canada, Guy et al. (2004) found 9 genotype B isolates, 3 genotype A isolates and 3 mixed (genotype A + genotype B). Results reported in our study agree with those from studies in Bangladesh published by Ng et al. (2005). They found 32 genotype B and 3 genotype A isolates. Results from all studies are not strictly comparable since amplifications were done on different G. lamblia genes and, in some of them, the populations were different, i.e., they were limited to symptomatic individuals. Genotype AII was found only in children and genotype B in both adults and children, although this difference cannot be considered significant. As regards sex, the distribution frequency of the genotypes was similar in both. In the animals studied, genotype B was detected in only one canine sample. However, a more detailed study about the zoonotic transmission of this parasitosis should include more samples as well as the analysis of dog genotypes (C, D) and livestock genotype (E). Thompson (2000) stated that in rural environments the transmission of dog genotypes occurs more frequently than in urban environments. In his study, the dog had the zoonotic genotype and lived in a rural area. In studies of G. lamblia isolated from cattle, genotype E was detected with a prevalence between 56 and 57% (O' Handley et al. 2000). This genotype has no zoonotic risk (Matsubayashi et al. 2005). In this study, there was no amplification from cattle feces. Overcrowding was present in three people infected with genotype AII, that enabled us to hypothesize the possibility of inter-human transmission. Most people with genotype B showed no overcrowding although house flooding was frequent, suggesting the possibility of transmission from a contaminated source such as water or food. Three people with genotype-AII G. lamblia reported appetite loss and weakness. None of them exhibited diarrhea, abdominal pain, sleeping disorders or vomiting, symptoms that were found in people with genotype-B. Only two people with genotype B reported no signs/symptoms. The results of this study correlated genotype AII with oligosymptomatic giardiasis profiles and genotype B with asyntomatic/polysyntomatic profiles. People infected with genotype B are likely to seek medical advice while those with genotype AII are not, resulting in slow deterioration of their own health status, due to alterations in the nutrient absorption caused by G. lamblia (Read et al. 2002). Amar et al. (2002) determined that genotype B was responsible for a diarrheal outbreak in a daycare center in England. However, a longitudinal study carried out in Australia by Read et al. (2002) reported that children infected with genotype A had a 26-fold greater risk of having diarrhea. In the Netherlands, Homan and Mank (2001) found that genotype B infection was associated with chronic diarrhea, while genotype A infections were related to intermittent diarrhea. Ceu Souza and Poiares da Silva (2004) and Eligio García et al. (2002) found no correlation between digestive manifestations and genotypes. Our study included people with and without symptoms since we hypothesized that selecting symptomatic patients only would not have shown the true prevalence of G. lamblia and the epidemiological role of the different genotypes. Oligo/asymptomatic people continue to attend school, the daycare center and workplace, producing localized endemic focuses. In conclusion, genotype AII showed a low prevalence and was detected in three children who lived together in a poor home in LP. The children were oligosymptomatic and none had diarrhea. This genotype was not found in animals. Genotype B showed a high prevalence in both adults and children. It was found in polysymptomatic people, many of whom presented diarrhea. It was found in only one animal (dog). Transmission may have occurred through a common source of infection since it was found in people from different homes sharing the same water supply and experiencing house flooding. Studies carried out with a higher number of samples, both human and animal, coming from different areas will increase our understanding of the epidemiology of giardiasis in Argentina. ACKNOWLEDGEMENT To Laura Cipolla for her help in the translator of the manuscript. REFERENCES
Copyright 2008 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08015t1.jpg] [oc08015t2.jpg] [oc08015f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}