 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 103, No. 2, March, 2008 , pp. 172-179 Preliminary characterization of a Rhodnius prolixus hemolymph trypanolytic protein, this being a determinant of Trypanosoma rangeli KP1(+) and KP1(-) subpopulations' vectorial ability Ximena Carolina Pulido, Gerardo Pérez1, Gustavo Adolfo Vallejo/+ Laboratorio de Investigaciones
en Parasitología Tropical, Facultad de Ciencias, Universidad
del Tolima, A.A. 546, Ibagué, Colombia 1Grupo
de Investigación en Proteínas, Departamento de Química,
Universidad Nacional de Colombia, Bogotá, Colombia Finnancial support: Instituto Colombiano para el Desarrollo de la Ciencia y la Tecnología Francisco José de Caldas (COLCIENCIAS) grant 1105-05-16919, and Universidad del Tolima, Colombia, Research Fund. Received 11 September
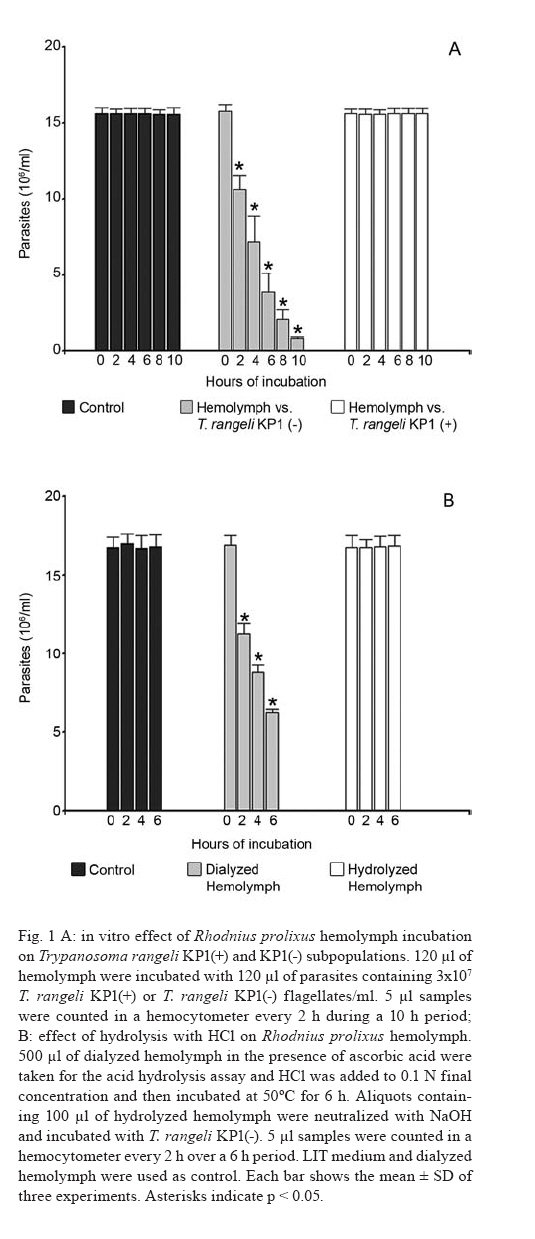
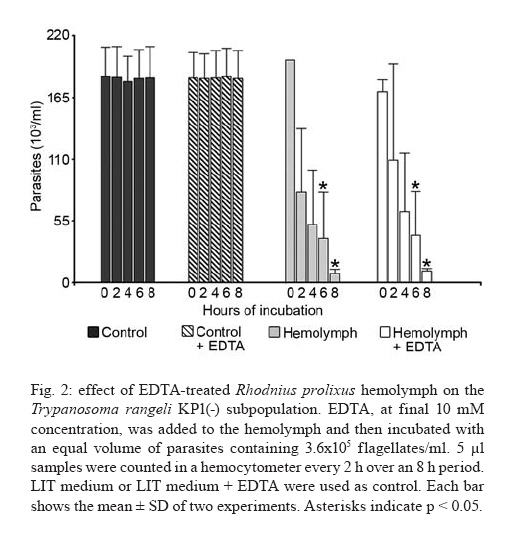
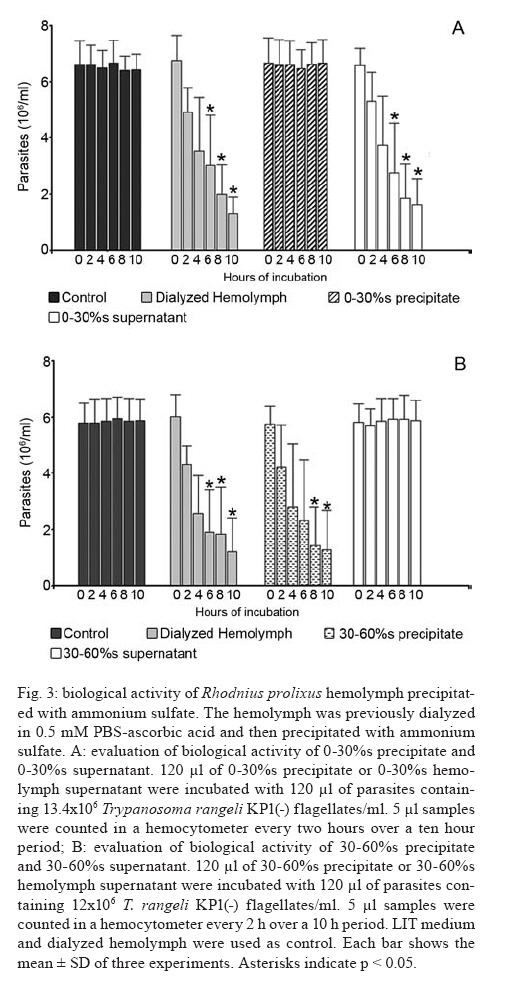
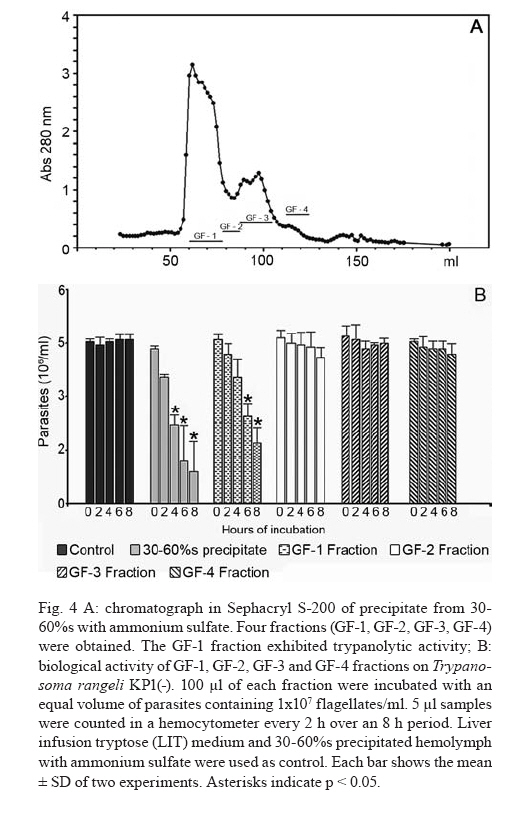
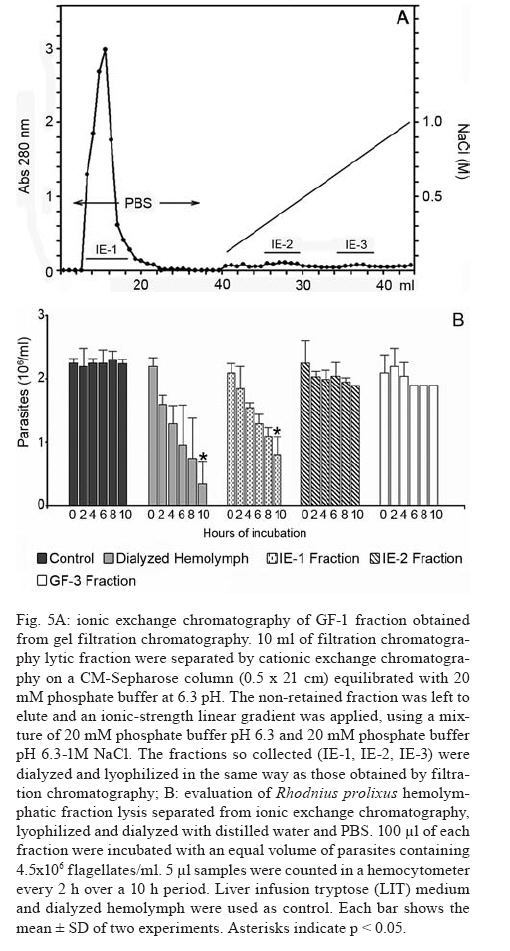
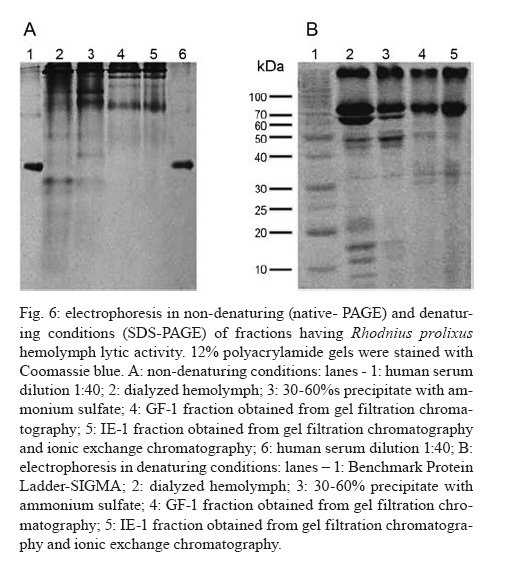
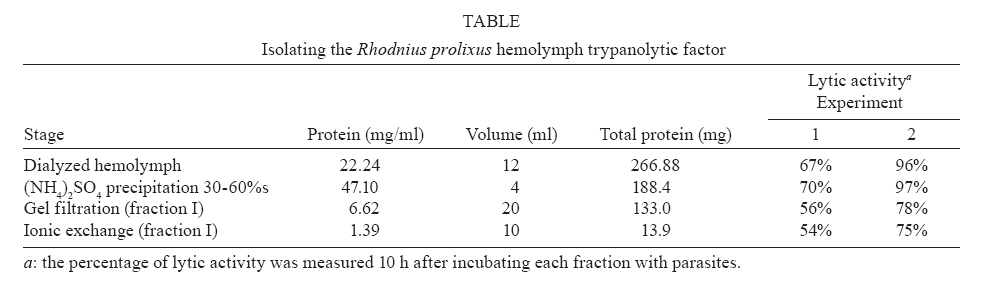
2007 Code Number: oc08029 Rhodnius prolixus is the main Trypanosoma rangeli vector in several Latin-American countries and is susceptible to infection with KP1(+) strains; however, it presents an invasion-resistant response to KP1(-) strains. The present work has identified a trypanolytic protein against T. rangeli KP1(-) in the R. prolixus hemolymph which was fractioned with ammonium sulfate (following dialysis). The results revealed a protein component which did not depend on divalent cations for its biological function whilst keeping its trypanolytic activity at temperatures ranging from -20ºC to 37ºC, at 7.0 to 10.5 pH. The protein was partially purified by gel filtration chromatography and ionic exchange chromatography. The major component presented a molecular weight of around 79 kDa and an isoelectric point between 4.9 and 6.3 and may be directly related to hemolymph trypanolytic activity against T. rangeli KP1(-) populations. Key words: Rhodnius prolixus - Trypanosoma rangeli KP1(+) - Trypanosoma rangeli KP1(-) - trypanolytic factor - vectorial ability - insect immunity Trypanosoma rangeli is a hemoflagellate which is apparently apathogenous for humans, presenting pathogenicity for the invertebrate host. T. rangeli is of great interest due to its presence in mixed infection with Trypanosoma cruzi in vertebrates and invertebrate hosts, thereby interfering with a clear diagnosis of Chagas' disease and the epidemiology of these parasites in different Latin-American countries (Guhl & Vallejo 2003). T. rangeli vectors become infected by ingesting blood from wild or domestic reservoirs in natural conditions. The parasite develops in the digestive tract and then invades the hemolymph where it is recognized by the vector's immune system; however, some strains may escape host defense mechanisms, reaching the salivary glands where they become infective forms or metacyclic trypomastigotes (D'Alessandro-Bacigalupo & Gore-Saravia 1992). Salivary gland invasion is a necessary step for transmission, being mediated by one or several ligand-receptor interactions (Basseri et al. 2002). Previous work has revealed two phylogenetically different groups in T. rangeli called T. rangeli KP1(-) and T. rangeli KP1(+), these being mainly transmitted by the 16 Rhodnius species described so far (Vallejo et al. 2002, 2007). Specificity between the vector and the parasite strain which it transmits has been pointed out; each Rhodnius species seems to select the T. rangeli subpopulation to which it is susceptible in nature so that it can then be transmitted to the vertebrate host via insect bite (Vallejo et al. 2003). Nevertheless, a close association has been observed between T. rangeli KP1(+) and vectors from the prolixus group (which includes Rhodnius prolixus and Rhodnius neglectus) and between T. rangeli KP1(-) and vectors from the pallescens group (including Rhodnius pallescens, Rhodnius colombiensis and Rhodnius ecuadoriensis) (Vallejo et al. 2003, Urrea et al. 2005). Recent parasite-vector interaction studies have shown a trypanolytic factor in R. prolixus hemolymph impeding the development of T. rangeli KP1(-) strains isolated from R. pallescens, R. colombiensis and R. ecuadoriensis (Sánchez et al. 2005). It has been stated that immune peptide and protein production in malaria, leishmaniasis and trypanosomiasis vectors could play an important role in limiting parasite infection (Boulanger et al. 2001, 2002a, 2004); this fact could explain different aspects related to these parasitosis' vectorial ability (Boulanger et al. 2002b, 2006). Most studies carried out on different vectors have led to identifying inducible peptides and proteins as a response to parasite infections. For example, gambycin is an antimicrobial peptide which has been isolated from the Anopheles vector; it is induced during early and late stages of malaria and is lethal for Plasmodium berghei ookinetes (Vizioli et al. 2001). A Phlebotomus duboscqui defensin has exhibited anti-parasite activity, specifically against Leishmania major promastigote forms (Boulanger et al. 2004). A complement system-like protein (TEP-1) from the Anopheles gambiae mosquito has promoted destruction of P. berghei stages in the insect's mid-gut (Blandin et al. 2004). An attacin isolated from Glossina morsitans morsitans has presented trypanolytic activity against Trypanosoma brucei blood-stage forms from mammals and stages developed inside the insect (Hao et al. 2003, Hu & Aksoy 2005). Other peptides or proteins have also been found which are constitutively present in the hemolymph or different insect secretions, such as an antimicrobial peptide "stomoxyn" detected in Stomoxis calcitrans presenting lytic activity against T. brucei rhodesiense (Boulanger et al. 2002a, b). A lytic protein called "trialysin" has been detected in Triatoma infestans saliva which acts on T. cruzi trypomastigotes (Amino et al. 2002). An antimicrobial peptide belonging to the family of small cationic peptides associated with innate immunity has been isolated from the R. prolixus hemolymph (López et al. 2003). The R. prolixus hemolymph trypanolytic factor was analyzed with the aim of contributing towards knowledge of factors regulating T. rangeli subpopulation transmission by different Rhodnius species. The effect of its digestion with pepsin and HCl for determining its protein nature was studied; the effect of variables such as adding chelate agents, temperature and pH were analyzed. This protein (which is probably implicated in T. rangeli KP1(-) strains' lysis) was also partially purified by gel filtration chromatography and ionic exchange chromatography. MATERIALS AND METHODS T. rangeli strains - The T. rangeli KP1(-) I/COL/CO/98/P53 strain isolated from the salivary glands of R. colombiensis from Coyaima (Tolima, Colombia) and T. rangeli KP1(+) I/PRX/CO/84/P19 strain isolated from the salivary glands of R. prolixus from Tibú (Norte de Santander, Colombia) were used in this work. Parasites were cultured in NNN medium supplemented with liver infusion tryptose (LIT), incubated at 28ºC; culture-Rhodnius-mouse cyclic passes were carried out each three months, according to the methodology described by Vallejo et al. (1986). Collecting R. prolixus hemolymph - 1,200 R. prolixus fifth-stage nymphs and adults were fed with chicken's blood; seven days after being fed, the hemolymph was extracted and centrifuged at 14,000 g for 3 min. The supernatant was dialyzed with 20 mM phosphate buffer pH 7.2, 150 mM NaCl (PBS) plus 0.5 mM ascorbic acid for controlling melanization. Incubating the R. prolixus hemolymph with T. rangeli KP1(-) and KP1(+) forms - 120 µl of R. prolixus hemolymph were incubated in a humid chamber with 120 µl of parasites suspended in LIT medium at 3x107parasites/ml concentration for 10 h. Parasites were incubated in the same conditions with LIT medium or heat-inactivated hemolymph as negative control. The parasites were counted in a Neubauer chamber every 2 h for 10 h. The experiment was repeated ten times, producing very close similar results. Digesting R. prolixus hemolymph with pepsin and acid hydrolysis - 6 µl pepsin (1 mg/ml) at pH 2.0 were mixed with 20 µl hemolymph and incubated at 37ºC in a water-bath for 4 h, followed by neutralization with concentrated NaOH. The solution was incubated with an equal volume of parasites; flagellates were counted two hours later. 500-600 µl of dialyzed hemolymph (in the presence of ascorbic acid) were taken for the acid hydrolysis assay; HCl was added to provide 0.1 N final concentration and then incubated at 50ºC for 24 h. Aliquots containing 100 µl of hydrolyzed hemolymph were taken at 0, 6, 10 and 24 h, neutralized with NaOH and incubated with T. rangeli KP1 (-). The experiment was repeated three times, producing similar results. Evaluating the effect of temperature, pH and adding chelate agents on R. prolixus hemolymph lytic activity - The hemolymph was kept at -20ºC and 5ºC during 24 h or it was incubated at 37ºC, 70ºC and 92ºC for 1 h. Aliquots containing the P53 strain were then incubated for 2 h with the treated hemolymph; the parasites were then counted.The effect of pH on hemolymph stability was evaluated using PBS pH 7.0, PBS + 5 mM thiourea pH 7.0 and 50 mM NaHCO3 pH 10.5; 20 µl parasites suspended in LIT medium were incubated with 10 µl dialyzed hemolymph and 10 µl of each buffer or 20 µl of each buffer or LIT medium. Parasites were counted 2 h later. Adding EDTA (10 mM or 50 mM final concentration) or EGTA (5 mM or 100 mM final concentration) to the hemolymph was evaluated; the biological test was carried out with the P53 strain. The experiment was repeated four times, leading to similar results. Isolating the R. prolixus hemolymph lytic fraction - Fractioned precipitation was carried out with (NH4)2SO4 on hemolymph dialyzed in 0.5 mM PBS-ascorbic acid at 0-30%s and 30-60%s. Precipitate having lytic activity was dissolved in 12 ml PBS and separated by filtration chromatography on a Sephacryl S-200 column (1.5 x 90 cm) equilibrated with PBS. Fractions were then grouped, each being dialyzed once with distilled water for 24 h and twice with 20 mM NH4HCO3 and used for electrophoresis or dialyzed in PBS and used for biological tests. Dialyzed samples were lyophilized and suspended in water for the biological assay with the parasites. Protein concentration was determined by Bradford's method (1976). 10 ml of filtration chromatography lytic fraction were separated by cationic exchange chromatography on a CM-Sepharose column (0.5 x 21cm) equilibrated with 20 mM phosphate buffer at 6.3 pH. The non-retained fraction was left to elute; an ionic-strength linear gradient was applied using a mixture of 20 mM phosphate buffer pH 6.3 and 1M NaCl. The fractions so collected were dialyzed and lyophilized in the same way as those obtained by filtration chromatography. Different fractions were submitted to electrophoresis in denaturing (SDS-PAGE) and non-denaturing conditions (native_PAGE), according to the methodology described by Laemmli (1970), using 4% concentration gel and 12% resolution gel. The molecular weight (MW) of the protein's major trypanolytic fraction component was determined by using standard MW markers (Benchmark Protein Ladder, SIGMA). Statistical analysis - The results were analyzed by ANOVA, Tukey's and LSD (least significant difference) tests, using OpenStat statistical software, version 2007 (www.statpages.org/miller/openstat), and InfoStat (2004). Differences between treatment and controls were not considered to be statistically significant (p > 0.05). RESULTS R. prolixus hemolymph in vitro incubation with T. rangeli KP1(+) and KP1(-) - Once 10 h had elapsed (following R. prolixus hemolymph being incubated with the T. rangeli KP1(-) P53 strain), it was observed that the number of parasites had significantly decreased whilst remaining constant in the control or when the hemolymph was incubated with the T. rangeli KP1(+) P19 strain (Fig. 1A). The number of KP1(-) parasites was statistically significant (p < 0.05) in the three experiments from two up to 10 h incubation when compared to control and the KP1(+) parasites. The nature of the trypanolytic factor - Pepsin-treated hemolymph completely eliminated parasite lytic activity (data not shown); hemolymph treated with acid hydrolysis for 6 h did not produce lytic activity (Fig. 1B). There were no significant differences (p > 0.05) between the number of parasites incubated with hydrolyzed hemolymph and the control at any time, thereby indicating the trypanolytic factor's protein nature. The effect of temperature, pH and chelate agents on R. prolixus hemolymph lytic activity - Thermal denaturing was most notable in hemolymph kept at 92ºC for 1 h, presenting lytic activity inhibition compared to hemolymph kept at 70ºC where 24% lysis on T. rangeli KP1(-) was recorded. The lytic effect was conserved in hemolymph kept at -20ºC and 5ºC for 24 h and was also observed in hemolymph kept at 28ºC and 37ºC for 1 h (data not shown). A preliminary experiment had shown that the P53 strain was more tolerant to PBS buffer pH 7.0 than PBS-thiourea pH 7.0 or carbonate pH 10.5. Once these buffers had been evaluated, it was found that hemolymph incubated with such buffers presented similar lytic activity as they exhibited 48.4%, 52% and 46.3% lysis, respectively, after 2 h incubation. These results showed that R. prolixus hemolymph lytic activity remained stable from 7 to 10.5 pH (data not shown). The lytic activity of hemolymph treated with EDTA was not seen to be affected when using the P53 strain, indicating that it was not divalent cation-dependent (Fig. 2). The number of KP1(-) parasites was not statistically significant (p > 0.05) in experiments between EDTA-treated hemolymph and non treated hemolymph at any time. The results obtained in the presence of EGTA were practically the same as those obtained with EDTA (data not shown), meaning that factor activity did not particularly depend on Ca+2. Isolating the lytic fraction in R. prolixus hemolymph - Fractioned precipitation assays with ammonium sulfate carried out with R. prolixus hemolymph showed that trypanolytic activity was present in the supernatant from 0-30%s and in precipitate from 30-60%s (Fig. 3A,B). The number of KP1(-) parasites was not statistically significant (p > 0.05) in experiments using dialyzed hemolymph and 0-30%s supernatant (Fig. 3A) or dialyzed hemolymph and 30-60%s precipitate (Fig. 3B). There were significant differences (p < 0.05) between the above mentioned lytic fractions and the control or 0-30%s precipitate and 30-60%s supernatant. Four fractions were obtained from gel filtration (GF) chromatography using the 30-60%s precipitate (Fig. 4A). The GF-1 fraction exhibited greater absorbance at 280 nm regarding the other fractions. This value was partly due to the protein concentration present (6.62 mg/ml) and partly to the presence of a yellow pigment, possibly presenting aromatic compounds, which also become absorbed in UV. The GF-1 fraction was characterized by presenting marked lytic activity on T. rangeli KP1(-) compared to GF-2, GF-3 and GF-4 fractions which were totally inactive, presenting similar behavior to that of the control (Fig. 4B) and the heat-inactivated GF-1 fraction (data not shown). Both the GF-1 fraction and precipitate having 30-60% saturation presented similar lytic behavior and significant differences (p < 0.05) at 4, 6 and 8 h with the control and GF-2, GF-3, GF-4 fractions (Fig. 4B). In another biological assay, once the GF-1 fraction had been inactivated by heat (80 ºC, 2 h), lysis became reduced to 9.3% compared to the non-inactivated GF-1 fraction and precipitate having 30-60% saturation, presenting 55.8% and 95% lytic activity, respectively (data not shown). These results revealed the presence of trypanolytic factor in the GF-1 fraction and confirmed its protein nature. Cationic exchange chromatography of the GF-1 fraction revealed elution with equilibrium buffer of a first fraction (IE-1) having high absorbance at 280nm, indicating that proteins present in this fraction were acidic. Two fractions were observed (IE-2, IE-3) when applying the ionic strength linear gradient, having very low absorbance values (Fig. 5A). The IE-1 fraction presented similar lytic activity when compared to dialyzed hemolymph; it had a significant difference (p < 0.05) after 10 h incubation when compared to control and IE-2 and IE-3 fractions (Fig. 5B). Electrophoresis in non-denaturing (native- PAGE) and denaturing conditions (SDS-PAGE) - Agreater 79 kDa band, together with five lesser bands having MW ranging from 30 kDa to 48 kDa, were observed in the GF-1 fraction in an electrophoretic run obtained in denaturing conditions (Fig. 6B). The GF-1 fraction was also separated into five protein bands having MW greater than 68.5 kDa in non-denaturing conditions (Fig. 6A). The presence of a band in the interface between concentration and separation gels was evident in all samples, probably due to protein aggregates. Analyzing the IE-1 fraction by SDS-PAGE (Fig. 6B) confirmed the presence of a major 79 kDa band. Table summarizes the results obtained in two experiments aimed at isolating the R. prolixus hemolymph trypanolytic factor. It should be noted that the differences observed in lysis percentages in both experiments were possibly due to changes in the proportion of short and long T. rangeli KP1(-) forms, even though the parasites used were always re-cultured each two days. A lesser lytic effect was consistently recorded in the first experiment than in the second one, due to long epimastigotes predominating over short ones; the long forms were thus more resistant to lysis. Lytic activity was more marked (96% and 75%) in the second experiment, since short forms predominated, these being more susceptible to trypanolytic factor action. DISCUSSION The epidemiological context of parasitosis demands that interactions between the parasite, invertebrate host and vertebrate host be fully understood. Parasite and vector co-evolution has involved physiological or biochemical strategies, facilitating or hampering parasite development in the invertebrate host (Azambuja et al. 2005). It has been observed that invertebrates have developed efficient immune mechanisms against parasite infections by producing potent antimicrobial peptides and polypeptides (Bulet et al. 2002). The first report about anti-trypanosome factors in triatomid hemolymph was published by Alvarenga and Bronfen (1982) who observed T. cruzi's inability to evolve in the triatomid hemocele. Another report by Azambuja et al. (1989) noted these factors in R. prolixus hemolymph against T. cruzi and other tripanosomatids. It should be stressed that R. prolixus infection with T. rangeli constitutes a very relevant natural model for studying parasite/vector interaction as most anti-parasite activity studies have been carried out on heterologous systems; molecules such as magainin isolated from frogs and cecropin isolated from silkworms have been proven against Plasmodium (Gwadz et al. 1989), cecropin isolated from Drosophila against Trypanosoma (Durvasula et al. 1997), insect defensins against Plasmodium (Shahabuddin et al. 1998), cecropin from Drosophila against Leishmania (Akuffo et al. 1998), diptericin isolated from Phormia against Trypanosoma sp. (Hao et al. 2001) and stomoxyn isolated from S. calcitrans against T. brucei rhodesiense (Boulanger et al. 2002b). Differing from the above studies, the present work has shown (for the first time) the existence of a protein which is constitutively present in R. prolixus hemolymph, behaving as a biological barrier and thereby determining the insect's vectorial ability. This molecule is responsible for inhibiting the development of T. rangeli KP1(-) forms and the insect is thus resistant to this parasite subpopulation. Differential behavior occurs when R. prolixus becomes naturally or experimentally infected with T. rangeli KP1(+) since the protein does not recognize this subpopulation and the parasite could then become transmitted to the vertebrate host by inoculation route (Sánchez et al. 2005, Vallejo et al. 2007). Ascorbic acid is an effective antioxidant as it reduces or prevents H2O2-induced peroxidation and the formation of free radicals (Frei et al. 1989, Deutsch 1998). Its use at low concentrations (0.5 mM) has substantially reduced oxidation in R. prolixus hemolymph and facilitated isolating the lytic factor in the present work. Acid hydrolysis and digestion with pepsin have been used in this work for determining the trypanolytic factor's protein nature, this being corroborated by hemolymph incubation at 70ºC and 92ºC for 1 h as lytic activity became reduced until becoming completely lost at 92ºC. The R. prolixus trypanolytic factor's biological activity remained stable at room temperature during parasite counting (10 h) or at 7 to 10.5 pH, possibly due to the protein's chemical or physical stability not being affected, thereby conserving its native state. Lytic activity against T. rangeli KP1(-) continued in EGTA and EDTA chelate agent-treated hemolymph; on the other hand, we deduced that the trypanolytic factor did not belong to the metalloprotein family or any lectin requiring divalent cations for carrying out its normal biological activity as described by Ashida et al. (1990), Feder et al. (1999), Yang et al. (2000), Konozy et al. (2003), Liu et al. (2006). Fractioned precipitation with ammonium sulfate (when isolating the lytic factor) led to total precipitation of the protein responsible for lytic activity in the 30-60%s range. This agent was used in the same saturation range during the initial stage of purifying a serpin six protease (a protein participating in activating immune mechanisms) from Manduca sexta hemolymph (Wang & Jiang 2004). GF chromatography led to eliminating low MW proteins which did not have any activity and obtaining a more active protein having a major 79 kDa MW component in the GF-1 fraction. Cationic exchange chromatography, using 20 mM phosphate buffer pH 6.3 as mobile phase, did not retain the protein and, consequently, the protein was negatively charged at this pH, meaning that it could be deduced that its isoelectric point (pI) was less than 6.3. Electrophoresis results in non-denaturing conditions revealed that lytic protein had less negative charge density than human seric albumin whose pI is 4.9. Bearing the lytic factor (probably 79 kDa) and seric albumin (68.5 kDa) MW in mind, it was deduced that its pI must be greater than 4.9. It is worth noting the trypanolytic factor's stability during the entire isolation procedure, as 75% activity was obtained at the end of such procedure. In short, we have considered that the major 79 kDa component was probably related to hemolymph trypanolytic activity against T. rangeli KP1(-) because this band was clearly observed in 30-60%s precipitate, and it was observed in GF-1 and IE-1 fractions which had lytic activity on parasites (Fig. 6B). However, the lytic nature of this 79 kDa component must be completely proven in further experiments. Another most important aspect was the trypanolytic factor's differential lysis (as observed by us) as it destroyed short epimastigote forms more rapidly than long ones which remained longer in in vitro biological tests (Table). These results agreed with other studies related to R. prolixus immune mechanisms which have demonstrated different behavior for short and long forms, as stated by Mello et al. (1999), who have shown that in vitro incubation of a purified R. prolixus hemolymph lectin with T. rangeli affected short epimastigote motility to a greater extent than that of long epimastigotes, and by Gomes et al. (1999), who found that T. rangeli short forms were more efficient when the R. prolixus prophenoloxidase system became activated than this parasite's long forms in in vivo and in vitro experiments. T. rangeli could be recognized in the vector insect's hemolymph and activate its defense system by cellular and humoral mechanisms which have been investigated in R. prolixus-T. rangeli interaction, amongst them being lysozymes and trypanolytic activity (Mello et al. 1995, Gomes et al. 1999), prophenoloxidase system activation (Mello et al. 1995, Gomes et al. 1999, Garcia et al. 2004), phagocytosis and hemocyte micro-agglutination (Mello et al. 1995, Garcia et al. 2004) hemolymph agglutination and nitric superoxide and oxide production (Whitten et al. 2001). It may be that all the elements forming part of the R. prolixus immune system interact together with the trypanolytic factor in in vivo conditions; the parasite thus disappears more rapidly in the hemolymph in natural conditions, happening 2 h post-infection as observed in several experiments. By contrast, lytic activity was a slower process in in vitro experiments, requiring at least 10 h for the parasites to disappear. It can thus be demonstrated that the R. prolixus-T. rangeli interaction is complex, since several factors intervene in it which are present in both the hemolymph's cellular and soluble fractions, thereby possibly making an integrated contribution towards eliminating T. rangeli KP1(-) strains. The trypanolytic factor described here explains the T. rangeli subpopulations' predominance in different parts of Colombia and other Latin-American countries. T. rangeli KP1(+) strains have been isolated from domiciliated R. prolixus in Honduras, Colombia and Venezuela, from wild R. neglectus in the Federal District of Brazil (Urrea et al. 2005) and wild R. prolixus in Casanare (Colombia) (unpublished data). On the other hand, T. rangeli KP1(-) strains have been isolated from wild R. pallescens from Panama and Colombia, from wild R. colombiensis in the central region of Colombia and from domiciliated R. ecuadoriensis in Peru (Vallejo et al. 2007). This body of work has led to concluding that T. rangeli subpopulation transmission specificity does not depend on a domestic or wild setting, but rather on the Rhodnius species possessing the factors promoting or limiting parasite infection. The mechanisms inhibiting T. rangeli KP1(+) transmission in R. pallescens, R. colombiensis and R. ecuadoriensis vectors remain unknown. R. prolixus-T. rangeli interaction is an important model which may be explored in other vectors' interaction with micro-organisms or parasites for isolating and identifying possible transmission-limiting factors, as these molecules could be used for controlling vector-transmitted diseases in the mid-term, such as Chagas' disease, leishmaniasis, malaria and filariasis. ACKNOWLEDGEMENTS To Dr. Felipe Guhl from the Centro de Investigaciones en Microbiología y Parasitología Tropical (CIMPAT), Universidad de los Andes, Bogotá, who kindly provided the T. rangeli I/PRX/CO/84/P19 strain. To Dr. Jairo Alfonso Clavijo from the Mathematics and Statistics Department, Universidad del Tolima for helping with the statistical analysis, Daniel Alfonso Urrea, provided helpful assistance with graphical designs. To the Universidad Nacional de Colombia's Chemistry Department for making its infrastructure and materials available for carrying out this work. To Jason Garry for patiently reading and correcting the manuscript. REFERENCES
Copyright 2008 - Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc08029f2.jpg] [oc08029f6.jpg] [oc08029f5.jpg] [oc08029t1.jpg] [oc08029f4.jpg] [oc08029f3.jpg] [oc08029f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}